Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Kwang Seok Ahn | + 2706 word(s) | 2706 | 2021-06-24 05:07:09 | | | |
| 2 | Vivi Li | Meta information modification | 2706 | 2021-07-06 09:03:17 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ahn, K.S. Resveratrol and TGF-β Signaling Pathway. Encyclopedia. Available online: https://encyclopedia.pub/entry/11705 (accessed on 08 August 2026).
Ahn KS. Resveratrol and TGF-β Signaling Pathway. Encyclopedia. Available at: https://encyclopedia.pub/entry/11705. Accessed August 08, 2026.
Ahn, Kwang Seok. "Resveratrol and TGF-β Signaling Pathway" Encyclopedia, https://encyclopedia.pub/entry/11705 (accessed August 08, 2026).
Ahn, K.S. (2021, July 06). Resveratrol and TGF-β Signaling Pathway. In Encyclopedia. https://encyclopedia.pub/entry/11705
Ahn, Kwang Seok. "Resveratrol and TGF-β Signaling Pathway." Encyclopedia. Web. 06 July, 2021.
Copy Citation
Resveratrol (Res) is a well-known natural product that can exhibit important pharmacological activities such as antioxidant, anti-diabetes, anti-tumor, and anti-inflammatory. An evaluation of its therapeutic effects demonstrates that this naturally occurring bioactive compound can target different molecular pathways to exert its pharmacological actions. Transforming growth factor-beta (TGF-β) is an important molecular pathway that is capable of regulating different cellular mechanisms such as proliferation, migration, and angiogenesis. TGF-β has been reported to be involved in the development of disorders such as diabetes, cancer, inflammatory disorders, fibrosis, cardiovascular disorders, etc.
resveratrol
transforming growth factor-beta (TGF-β)
chronic diseases
fibrosis
cancer
diabetes
therapy
1. Resveratrol
From immemorial times, plant-derived natural compounds have been under attention in the treatment of different disorders such as inflammatory diseases, cancers, pulmonary diseases, metabolic disorders, neurological disorders (NDs) including Alzheimer’s disease (AD) and Parkinson’s disease (PD), infertility, and so on [1][2][3][4][5][6][7][8][9][10]. Phytochemicals can exhibit beneficial actions against diseases due to their excellent pharmacological activities [11][12][13][14]. These benefits have resulted in extensive research into finding new natural compounds and revealing their potential mechanisms of actions [15][16][17]. Resveratrol (Res) is a dietary phytochemical that has been reported to be efficacious treatment for various ailments by targeting diverse molecular pathways [18][19][20][21]. The role of Res in the treatment of chronic diseases was established in early 1990s when it was found that this phytochemical possesses significant cardioprotective benefits [22]. This ascending trend toward Res research led to the revelation of its significant biological and therapeutic activities. The first report about anti-tumor activity of Res dates back to 1997, when Jang and his colleagues reported its inhibitory effect on leukemia [23].
Currently, Res can be derived from various plants including Arachis hypogea, Cassia sp., Eucalyptus sp., Morus rubra, and so on using a number of different isolation techniques [24]. High-performance liquid chromatography is the best strategy [25][26][27][28]. Over the past decades, Res has been applied in the treatment of various diseases such as osteoarthritis [29][30][31], NDs [32], cancer [33][34][35], diabetes [36], cardiovascular diseases [37], liver disorders [38], and so on. An increasing amount of evidence is in agreement with the fact that Res affects different molecular pathways to exhibit its protective effects [39][40][41]. Hence, the identification of these targets can promote further studies for investigating molecular pathways and the mechanisms of its therapeutic actions in depth. For instance, anti-inflammation is one of the most important biological effects of Res treatment. To function as an anti-inflammatory molecule, Res can effectively inhibit the activation of pro-inflammatory transcription factors such as nuclear factor-kappaB (NF-ĸB). It seems that the anti-inflammatory actions of Res are not only mediated via inhibitory actions on the NF-ĸB signaling pathway, but they also rely on its action as a PARP-γ agonist [42]. The anti-inflammatory activities of Res are also characterized by decreased levels of interleukin (IL)-6, IL-8, and tumor necrosis factor-α (TNF-α), etc. [43]. The production of pro-inflammatory lipid mediators from arachidonic acid can be mediated by the cyclooxygenase (COX) pathway. A number of anti-inflammatory drugs have been developed based on their inhibitory effect on COX-1 and COX-2 [44][45]. Res is capable of binding to the active site of COX-1 and thus causing anti-inflammatory effects. In addition to targeting inflammation, Res attaches to the active site of COX-2 to suppress cancer proliferation [46][47][48][49]. It is noteworthy that the inhibitory effect of Res on COX has been noted to follow a dose-dependent kinetics [50].
Obesity is one of the challenges faced in today’s world. Res has demonstrated great potential in reducing weight and exerting anti-obesity activity. Res changes white adipose tissue (WAT) into brown adipose tissue (BAT), which in turn decreases weight and improves insulin resistance [51]. The inhibitory action of Res on lipid accumulation leads to its effect on cardiovascular disorders. Res stimulates PARP-α/γ to activate ATP binding cassette (ABC) transporter A1/G1-mediated cholesterol efflux, resulting in a decrease in lipid accumulation and cholesterol levels. These effects can lead to a significant amelioration of atherosclerosis [52]. Based on the effect of Res on amyloid-beta (Aβ), this plant-derived natural compound is of importance in treating NDs. For instance, Res is able to inhibit inflammation and the microglial activation caused by Aβ. This results in the alleviation of inflammation (down-regulation of TNF-α and IL-6) and a diminution in apoptosis (caspase-1 down-regulation) [53]. The antioxidant activity of Res provides its protective effect during kidney injury. In rats exposed to nicotine, an increase occurs in oxidative stress markers via the down-regulation of glutathione. The administration of Res has been also correlated with improving the antioxidant defense system that protects renal cells against oxidative injury [54]. A newly published study also demonstrates the effect of Res on stem cells. Res can stimulate stem cell function to ameliorate pancreatic injury such as fibrosis and apoptosis [55]. Overall, these reports exhibit that Res has diverse therapeutic effects that have resulted in its extensive application in the treatment of various disorders [56][57][58]. In the current review, we specifically focus on the therapeutic effects of Res mediated by its regulatory action on the transforming growth factor-β (TGF-β) signaling pathway.
2. Resveratrol and TGF-β Signaling Pathway
2.1. Resveratrol and Fibrosis
Pulmonary fibrosis (PF) is a common disorder of the lung that is characterized with hypoxemia, restrictive functional ventilatory disturbance, and chronic fibrosis. Clinical manifestations of PF include wheezing, difficulties in breathing, and dry coughs [59]. The pathogenesis of PF is still not completely understood, but it appears that the TGF-β signaling pathway plays a significant role in PF development [60]. Thus, the administration of Res may be an ideal strategy in the amelioration of PF, and different molecular pathways may be involved. Normally, microRNA (miR)-21 can induce PF via the activation of TGF-β signaling and providing Smad7 nuclear translocation. TGF-β provides a positive feedback loop, so TGF-β enhances the expression of miR-21 and AP-1. The administration of Res down-regulates the expression of miR-21 via inhibition of the MAPK/AP-1 axis. This leads to a diminution in TGF-β expression and inhibition of Smad7, resulting in the alleviation of PF [61]. Accumulating data demonstrate that during the inhibition of fibrosis, Res affects the TGF-β signaling pathway via the modulation of miRs. Myocardial fibrosis (MF) is caused by the accumulation of collagen fibers, enhanced collagen content, and alteration in collagen composition. Systolic and diastolic functions of the heart can be negatively affected by MF [62]. TGF-β is one of the key players regulating MF [63]. The TGF-β/Smad7 axis can also contribute to the development of MF. The administration of Res can up-regulate the expression of miR-17, which in turn remarkably reduces levels of Smad7, leading to an improvement in MF [64].
In addition to PF and MF, renal fibrosis (RF) can arise as a result of the activation of the TGF-β signaling pathway. It has been reported that the inhibition of the TGF-β signaling pathway by natural products such as bardoxolone and nimbolide is of importance in RF therapy [65][66]. It is worth mentioning that Res can target the TGF-β signaling pathway, thereby causing an amelioration of RF. In RF treatment, fibroblast–myofibroblast differentiation (FMD), EMT, and the proliferation of tubular epithelial cells (TECs) should be targeted. The administration of Res can disrupt Smad2/3 activation by TGF-β and consequently suppress the proliferation of TECs, FMD, and EMT [67]. Increasing evidence demonstrates that EMT may be involved in renal fibrogenesis, and its activation can facilitate the development of RF [68][69][70][71]. Res is capable of suppressing EMT-mediated RF. It seems that TGF-β1 functions as an upstream mediator of EMT, and Res suppresses EMT and RF through inhibiting TGF-β1 [72]. In fact, in the stimulation of anti-fibrotic activity, Res affects the proliferation and survival of fibroblasts. It has been shown that Res can stimulate apoptosis in fibroblasts and suppress their growth as well. An investigation of the molecular pathways demonstrates that in targeting fibroblasts, Res can suppress TGF-β and the Smad2/3/4 complex, and it can also upregulate Smad7 [73].
It is worth mentioning that the anti-fibrotic activity of Res is dose-dependent, and using low doses is preferred as compared to higher doses. An experiment has evaluated the role of dose in the anti-fibrotic activity of Res. TGF-β induces fibrosis via formation of the Smad3/4 complex and subsequent stimulation of EMT. The administration of Res has been correlated with the deacetylation of Smad3 and Smad4 via sirtuin 1 (SIRT1). According to in vitro results, low doses of Res (5–20 mM) effectively exerted anti-fibrotic activity, while high doses (more than 40 mM) did not demonstrate any substantial anti-fibrotic activity. The in vivo findings are in line with in vitro results, so that low doses of Res (less than 25 mg/kg) improve fibrosis, while high doses of Res (more than 50 mg/kg) deteriorated the condition [74]. This study confirms the dose-related toxicity of Res. Overall, these studies demonstrate that TGF-β can function as a key player in the development of fibrosis and Res can suppress the TGF-β signaling pathway and its downstream targets such as Smads to alleviate fibrosis [75][76].
The TGF-β signaling pathway contributes to the development of fibrosis in different vital organs of body such as the lung and heart. The interesting point to highlight is the possible epigenetic regulation of TGF-β by miRs in the development of fibrosis. Res is capable of suppressing miR and TGF-β interaction in fibrosis therapy. MiR-17 and miR-21 are two important miRs that contribute to the emergence of myocardial and pulmonary fibrosis via TGF-β induction. The regulation of TGF-β by miRs is suppressed upon Res administration. RF also occurs by the function of TGF-β and subsequent induction of EMT. The TGF-β/EMT axis is inhibited by Res to alleviate RF. It is noteworthy that in the amelioration of fibrosis, components of TGF-β signaling such as Smad7 and Smad4 can also be down-regulated. Therefore, TGF-β is a versatile agent in the amelioration of fibrosis.
2.2. Resveratrol and Cancer Therapy
Accumulating data exhibit that the TGF-β signaling pathway can regulate both the proliferation and metastasis of cancer cells, and its inhibition is a promising strategy in cancer therapy [77][78][79][80][81][82]. Metastasis is an increasing challenge in the effective treatment of cancer. Cancer cells are able to migrate into neighboring and distant tissues, demanding novel strategies in the inhibition of their metastasis. EMT is one of the mechanisms that can promote invasion via the transformation of static epithelial cells into migratory mesenchymal ones [83]. A number of different molecular pathways have been recognized as regulators of EMT [84][85], and it has been found that TGF-β is capable of elevating migration via EMT induction. In breast cancer, TGF-β can stimulate EMT via Smad2 and Smad3 activation, leading to an increase in N-cadherin and vimentin levels, and a decrease in E-cadherin levels. The administration of Res suppresses the metastasis of breast cancer (under both in vitro and in vivo conditions) via the inhibition of TGF-β1 and down-regulation of Smad2 and Smad3 [86]. TGF-β also contributes to the migration and malignant behavior of lung cancer. In addition to breast cancer, Res targets TGF-β to inhibit EMT in lung cancer. By suppressing levels of TGF-β, Res down-regulates the levels of vimentin and fibronectin, while it enhances E-cadherin levels, leading to an inhibition of EMT and metastasis of lung cancer cells [87]. It is noteworthy that EMT induction enhances viability via the stimulation of cancer stem cell markers such as Bmi1 and Sox2. By inhibition of the TGF-β/Smad axis, Res not only inhibits EMT and migration, but also interferes with the proliferation and survival of cancer cells [88]. So, Res can function as a potential modulator of EMT in cancer cells to negatively affect their proliferation and metastasis.
Accumulating data also show that Res is able to diminish levels of TGF-β that in turn, suppresses the development of renal carcinoma [89]. These studies are in agreement with the fact that the inhibition of TGF-β by Res is of interest in suppressing tumor growth and metastasis [90]. Moreover, a dual relationship has been found between TGF-β and programmed cell death-1 (PD-1). For instance, PD-1 overexpression is associated with the induction of TGF-β, and TGF-β can regulate PD-1 expression [91][92]. This dual relationship is of importance in cancer therapy. Res can suppress the proliferation of oral cancer cells via the down-regulation of TGF-β and subsequent inhibition of PD-1. L-thyroxine as a thyroid hormone can also modulate the anti-tumor activity of Res via regulating the TGF-β/PD-1 axis [88].
Overall, the regulation of TGF-β by Res in cancer is of importance in terms of suppressing both migration and proliferation. The most well-known mechanism targeted by TGF-β is EMT, which can promote cancer metastasis. In addition, TGF-β can activate the signaling pathways such as PD-1 and Sox2 to ensure the growth and survival of cancer cells. Upon Res administration, TGF-β and its downstream targets are inhibited to pave the road for effective cancer therapy.
2.3. Resveratrol and Lung Injury
Injuries to vascular endothelium and alveolar epithelium by inflammatory factors can lead to the emergence of acute lung injury (ALI) [93]. Infections are able to generate ALI and among them, Pseudomonas aerogenosa, Candidate albicans, and staphylococcal enterotoxin B (SEB) are of importance [94][95][96]. In the amelioration of SEB-mediated lung injury, Res can target the TGF-β signaling pathway. Res can down-regulate the expression of miR-193a to inhibit TGF-β2 and TGFβR3, thus resulting in a decrease in levels of inflammatory cytokines and T cell infiltration [97]. The enhanced level of TGF-β has been associated with the development of asthma and lung injury [98]. In fact, the administration of Res may alleviate lung injury and asthma via decreasing levels of TGF-β [99]. Chronic obstructive pulmonary disease (COPD) is one of the most common disorders of lung tissue. Cigarette smoking is the most well-known reason for COPD [100]. Pulmonary inflammation, airflow obstruction, and remodeling are features of COPD [101]. Chronic inflammation can result in the development of COPD, and TGF-β has been found to play an important role in the pathogenesis of this disease [102][103]. Therefore, based on the modulatory impact of Res on TGF-β, the administration of this naturally occurring compound can be advantageous in the amelioration of COPD. It was also found that Res can decrease fibrotic response and inhibit mucus hypersecretion via the down-regulation of TGF-β [104].
It seems that via the regulation of TGF-β, Res is capable of reducing inflammation in lung and preventing the development of pathological events such as ALI, COPD, and asthma. Interestingly, Res inhibits inflammation via reducing the infiltration of cytokines and T cells. COPD is also emerged via pulmonary inflammation and fibrosis. Based on the effect of Res on TGF-β and subsequent decrease in fibrotic response and mucus hypersecretion, it can be beneficial in the treatment of COPD.
2.4. Resveratrol and Brain Injury
Cerebral hemorrhage is a leading cause of brain injury and vasospasm [105]. This malignancy results in ischemic/reperfusion and the induction of apoptosis in cancer cells [106][107]. The TGF-β signaling pathway has been correlated with brain injury [108]. Interestingly, the administration of Res was found to improve the blood–brain barrier (BBB) and inhibit apoptosis in neuronal cells. These protective effects of Res were found to be mediated via the inhibition of TGF-β-mediated ERK [109]. Moreover, it was found that exposing rats to alcohol is associated with an increase in levels of cytokines such as TGF-β. An administration of Res (10 and 20 mg/kg) can significantly improve cognitive deficits and reduces brain injury via decreasing TGF-β levels [110]. So, the alleviation of cognitive deficits and maintaining the integrity of BBB are functions of Res that can be mediated by TGF-β modulation.
2.5. Resveratrol and DM
During DM, microvascular complications can lead to hyperglycemia that accounts for the emergence of diabetic nephropathy (DN). Interestingly, an enhanced level of oxidative stress, renal polyol formation, protein kinase C induction, and activation of AMPK as well as the accumulation of advanced glycation end-products (AGEs) are responsible for DN [111][112]. TGF-β1 is considered as one of the potential pathways involved in the emergence of DN [113]. A combination of Res and rosuvastatin (RSU) was found to be beneficial in the alleviation of DN via the down-regulation of TGF-β1 [114]. The in vivo studies have also indicated that the administration of Res is a promising strategy in alleviating DN. It was observed that Res could diminish urinary albumin excretion, glomerular hypertrophy, and the deposition of fibronectin and collagen type IV to ameliorate DN. Moreover, an investigation of molecular pathways demonstrated that Res can alleviate TGF-β expression as well as the phosphorylation of Smad2 and Smad3 for DN alleviation (Figure 2) [115]. The most important effect of Res during DN is reducing fibrosis, which can be mediated via TGF-β inhibition.
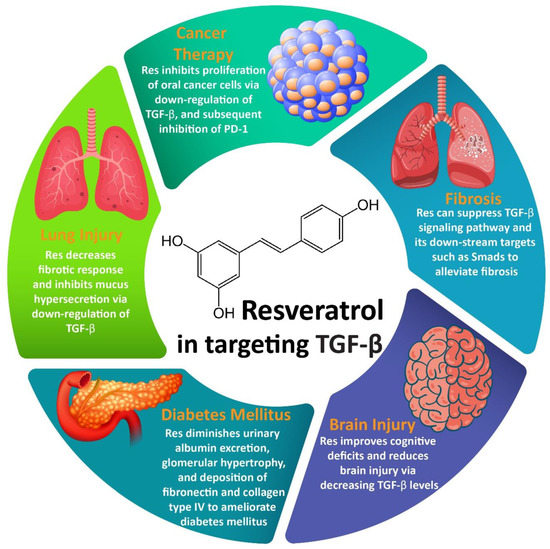
Figure 2. Regulation of TGF-β signaling by Res and its association with therapeutic effects.
References
- Mohan, C.D.; Rangappa, S.; Preetham, H.D.; Chandra Nayaka, S.; Gupta, V.K.; Basappa, S.; Sethi, G.; Rangappa, K.S. Targeting STAT3 signaling pathway in cancer by agents derived from Mother Nature. Semin. Cancer Biol. 2020.
- Aggarwal, V.; Tuli, H.S.; Thakral, F.; Singhal, P.; Aggarwal, D.; Srivastava, S.; Pandey, A.; Sak, K.; Varol, M.; Khan, M.A.; et al. Molecular mechanisms of action of hesperidin in cancer: Recent trends and advancements. Exp. Biol. Med. 2020, 245, 486–497.
- Baek, S.H.; Ko, J.H.; Lee, H.; Jung, J.; Kong, M.; Lee, J.W.; Lee, J.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; et al. Resveratrol inhibits STAT3 signaling pathway through the induction of SOCS-1: Role in apoptosis induction and radiosensitization in head and neck tumor cells. Phytomedicine Int. J. Phytother. Phytopharm. 2016, 23, 566–577.
- Dai, X.; Zhang, J.; Arfuso, F.; Chinnathambi, A.; Zayed, M.E.; Alharbi, S.A.; Kumar, A.P.; Ahn, K.S.; Sethi, G. Targeting TNF-related apoptosis-inducing ligand (TRAIL) receptor by natural products as a potential therapeutic approach for cancer therapy. Exp. Biol. Med. 2015, 240, 760–773.
- Prasannan, R.; Kalesh, K.A.; Shanmugam, M.K.; Nachiyappan, A.; Ramachandran, L.; Nguyen, A.H.; Kumar, A.P.; Lakshmanan, M.; Ahn, K.S.; Sethi, G. Key cell signaling pathways modulated by zerumbone: Role in the prevention and treatment of cancer. Biochem. Pharmacol. 2012, 15, 1268–1276.
- Tan, S.M.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of beta-escin as a novel inhibitor of signal transducer and activator of transcription 3/Janus-activated kinase 2 signaling pathway that suppresses proliferation and induces apoptosis in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2010, 334, 285–293.
- Lee, J.H.; Chinnathambi, A.; Alharbi, S.A.; Shair, O.H.M.; Sethi, G.; Ahn, K.S. Farnesol abrogates epithelial to mesenchymal transition process through regulating Akt/mTOR pathway. Pharmacol. Res. 2019, 150, 104504.
- Wong, A.L.A.; Hirpara, J.L.; Pervaiz, S.; Eu, J.Q.; Sethi, G.; Goh, B.C. Do STAT3 inhibitors have potential in the future for cancer therapy? Expert Opin. Investig. Drugs 2017, 26, 883–887.
- Ochiai, A.; Kuroda, K. Preconception resveratrol intake against infertility: Friend or foe? Reprod. Med. Biol. 2019, 19, 107–113.
- McSweeney, K.R.; Gadanec, L.K.; Qaradakhi, T.; Gammune, T.M.; Kubatka, P.; Caprnda, M.; Fedotova, J.; Radonak, J.; Kruzliak, P.; Zulli, A. Impridone enhances vascular relaxation via FOXO1 pathway. Clin. Exp. Pharmacol. Physiol. 2020.
- Kashyap, D.; Tuli, H.S.; Yerer, M.B.; Sharma, A.; Sak, K.; Srivastava, S.; Pandey, A.; Garg, V.K.; Sethi, G.; Bishayee, A. Natural product-based nanoformulations for cancer therapy: Opportunities and challenges. Semin. Cancer Biol. 2019.
- Shanmugam, M.K.; Manu, K.A.; Ong, T.H.; Ramachandran, L.; Surana, R.; Bist, P.; Lim, L.H.; Kumar, A.P.; Hui, K.M.; Sethi, G. Inhibition of CXCR4/CXCL12 signaling axis by ursolic acid leads to suppression of metastasis in transgenic adenocarcinoma of mouse prostate model. Int. J. Cancer 2011, 129, 1552–1563.
- Ramachandran, L.; Manu, K.A.; Shanmugam, M.K.; Li, F.; Siveen, K.S.; Vali, S.; Kapoor, S.; Abbasi, T.; Surana, R.; Smoot, D.T.; et al. Isorhamnetin inhibits proliferation and invasion and induces apoptosis through the modulation of peroxisome proliferator-activated receptor γ activation pathway in gastric cancer. J. Biol. Chem. 2012, 287, 38028–38040.
- Varughese, R.S.; Lam, W.S.-T.; Marican, A.A.b.H.; Viganeshwari, S.H.; Bhave, A.S.; Syn, N.L.; Wang, J.; Wong, A.L.-A.; Kumar, A.P.; Lobie, P.E.; et al. Biopharmacological considerations for accelerating drug development of deguelin, a rotenoid with potent chemotherapeutic and chemopreventive potential. Cancer 2019, 125, 1789–1798.
- Siveen, K.S.; Mustafa, N.; Li, F.; Kannaiyan, R.; Ahn, K.S.; Kumar, A.P.; Chng, W.J.; Sethi, G. Thymoquinone overcomes chemoresistance and enhances the anticancer effects of bortezomib through abrogation of NF-kappaB regulated gene products in multiple myeloma xenograft mouse model. Oncotarget 2014, 5, 634–648.
- Li, F.; Shanmugam, M.K.; Chen, L.; Chatterjee, S.; Basha, J.; Kumar, A.P.; Kundu, T.K.; Sethi, G. Garcinol, a polyisoprenylated benzophenone modulates multiple proinflammatory signaling cascades leading to the suppression of growth and survival of head and neck carcinoma. Cancer Prev. Res. 2013, 6, 843–854.
- Rajendran, P.; Li, F.; Shanmugam, M.K.; Vali, S.; Abbasi, T.; Kapoor, S.; Ahn, K.S.; Kumar, A.P.; Sethi, G. Honokiol inhibits signal transducer and activator of transcription-3 signaling, proliferation, and survival of hepatocellular carcinoma cells via the protein tyrosine phosphatase SHP-1. J. Cell. Physiol. 2012, 227, 2184–2195.
- Huang, X.-T.; Li, X.; Xie, M.-L.; Huang, Z.; Huang, Y.-X.; Wu, G.-X.; Peng, Z.-R.; Sun, Y.-N.; Ming, Q.-L.; Liu, Y.-X. Resveratrol: Review on its discovery, anti-leukemia effects and pharmacokinetics. Chem. Biol. Interact. 2019, 306, 29–38.
- Ko, J.-H.; Sethi, G.; Um, J.-Y.; Shanmugam, M.K.; Arfuso, F.; Kumar, A.P.; Bishayee, A.; Ahn, K.S. The Role of Resveratrol in Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2589.
- Shanmugam, M.K.; Warrier, S.; Kumar, A.P.; Sethi, G.; Arfuso, F. Potential Role of Natural Compounds as Anti-Angiogenic Agents in Cancer. Curr. Vasc. Pharmacol. 2017, 15, 503–519.
- Frazzi, R.; Guardi, M. Cellular and molecular targets of resveratrol on lymphoma and leukemia cells. Molecules 2017, 22, 885.
- Garg, A.K.; Buchholz, T.A.; Aggarwal, B.B. Chemosensitization and radiosensitization of tumors by plant polyphenols. Antioxid. Redox Signal. 2005, 7, 1630–1647.
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.; Fong, H.H.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 1997, 275, 218–220.
- Aggarwal, B.B.; Bhardwaj, A.; Aggarwal, R.S.; Seeram, N.P.; Shishodia, S.; Takada, Y. Role of resveratrol in prevention and therapy of cancer: Preclinical and clinical studies. Anticancer Res. 2004, 24, 2783–2840.
- Ragab, A.S.; Van Fleet, J.; Jankowski, B.; Park, J.-H.; Bobzin, S.C. Detection and quantitation of resveratrol in tomato fruit (Lycopersicon esculentum Mill.). J. Agric. Food Chem. 2006, 54, 7175–7179.
- Lo, C.; Le Blanc, J.Y.; Yu, C.K.; Sze, K.; Ng, D.C.; Chu, I.K. Detection, characterization, and quantification of resveratrol glycosides in transgenic arabidopsis over-expressing a sorghum stilbene synthase gene by liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. Int. J. Devoted Rapid Dissem. Minute Res. Mass Spectrom. 2007, 21, 4101–4108.
- Loizzo, M.R.; Nigro, S.; De Luca, D.; Menichini, F. Detection of ochratoxin A and cis-and trans-resveratrol in red wines and their musts from Calabria (Italy). Food Addit. Contam. Part A 2011, 28, 1561–1568.
- Koga, C.C.; Becraft, A.R.; Lee, Y.; Lee, S.Y. Taste detection thresholds of resveratrol. J. Food Sci. 2015, 80, S2064–S2070.
- Xu, X.; Liu, X.; Yang, Y.; He, J.; Gu, H.; Jiang, M.; Huang, Y.; Liu, X.; Liu, L. Resveratrol inhibits the development of obesity-related osteoarthritis via the TLR4 and PI3K/Akt signaling pathways. Connect. Tissue Res. 2019, 60, 571–582.
- Ebrahim, H.A.; Alzamil, N.M.; Al-Ani, B.; Haidara, M.A.; Kamar, S.S.; Dawood, A.F. Suppression of knee joint osteoarthritis induced secondary to type 2 diabetes mellitus in rats by resveratrol: Role of glycated haemoglobin and hyperlipidaemia and biomarkers of inflammation and oxidative stress. Arch. Physiol. Biochem. 2020, 1–8.
- Zhang, G.; Zhang, H.; You, W.; Tang, X.; Li, X.; Gong, Z. Therapeutic effect of Resveratrol in the treatment of osteoarthritis via the MALAT1/miR-9/NF-κB signaling pathway. Exp. Ther. Med. 2020, 19, 2343–2352.
- Cosín-Tomàs, M.; Senserrich, J.; Arumí-Planas, M.; Alquézar, C.; Pallàs, M.; Martín-Requero, Á.; Suñol, C.; Kaliman, P.; Sanfeliu, C. Role of Resveratrol and Selenium on Oxidative Stress and Expression of Antioxidant and Anti-Aging Genes in Immortalized Lymphocytes from Alzheimer’s Disease Patients. Nutrients 2019, 11, 1764.
- Yuan, L.; Zhou, M.; Huang, D.; Wasan, H.S.; Zhang, K.; Sun, L.; Huang, H.; Ma, S.; Shen, M.; Ruan, S. Resveratrol inhibits the invasion and metastasis of colon cancer through reversal of epithelial- mesenchymal transition via the AKT/GSK-3β/Snail signaling pathway. Mol. Med. Rep. 2019, 20, 2783–2795.
- Jang, Y.G.; Go, R.E.; Hwang, K.A.; Choi, K.C. Resveratrol inhibits DHT-induced progression of prostate cancer cell line through interfering with the AR and CXCR4 pathway. J. Steroid Biochem. Mol. Biol. 2019, 192, 105406.
- Kiskova, T.; Kubatka, P.; Büsselberg, D.; Kassayova, M. The Plant-Derived Compound Resveratrol in Brain Cancer: A Review. Biomolecules 2020, 10, 161.
- Rašković, A.; Ćućuz, V.; Torović, L.; Tomas, A.; Gojković-Bukarica, L.; Ćebović, T.; Milijašević, B.; Stilinović, N.; Cvejić Hogervorst, J. Resveratrol supplementation improves metabolic control in rats with induced hyperlipidemia and type 2 diabetes. Saudi Pharm. J. SPJ Off. Publ. Saudi Pharm. Soc. 2019, 27, 1036–1043.
- Hong, M.; Li, J.; Li, S.; Almutairi, M.M. Resveratrol Derivative, Trans-3, 5, 4′-Trimethoxystilbene, Prevents the Developing of Atherosclerotic Lesions and Attenuates Cholesterol Accumulation in Macrophage Foam Cells. Mol. Nutr. Food Res. 2020, 64, e1901115.
- Yu, B.; Qin, S.Y.; Hu, B.L.; Qin, Q.Y.; Jiang, H.X.; Luo, W. Resveratrol improves CCL4-induced liver fibrosis in mouse by upregulating endogenous IL-10 to reprogramme macrophages phenotype from M(LPS) to M(IL-4). Biomed. Pharmacother. 2019, 117, 109110.
- Tewari, D.; Nabavi, S.F.; Nabavi, S.M.; Sureda, A.; Farooqi, A.A.; Atanasov, A.G.; Vacca, R.A.; Sethi, G.; Bishayee, A. Targeting activator protein 1 signaling pathway by bioactive natural agents: Possible therapeutic strategy for cancer prevention and intervention. Pharm. Res. 2018, 128, 366–375.
- Deng, S.; Shanmugam, M.K.; Kumar, A.P.; Yap, C.T.; Sethi, G.; Bishayee, A. Targeting autophagy using natural compounds for cancer prevention and therapy. Cancer 2019, 125, 1228–1246.
- Mishra, S.; Verma, S.S.; Rai, V.; Awasthee, N.; Chava, S.; Hui, K.M.; Kumar, A.P.; Challagundla, K.B.; Sethi, G.; Gupta, S.C. Long non-coding RNAs are emerging targets of phytochemicals for cancer and other chronic diseases. Cell. Mol. Life Sci. CMLS 2019, 76, 1947–1966.
- Ben Lagha, A.; Andrian, E.; Grenier, D. Resveratrol attenuates the pathogenic and inflammatory properties of Porphyromonas gingivalis. Mol. Oral Microbiol. 2019, 34, 118–130.
- Farzanegan, A.; Shokuhian, M.; Jafari, S.; Shirazi, F.S.; Shahidi, M. Anti-histaminic Effects of Resveratrol and Silymarin on Human Gingival Fibroblasts. Inflammation 2019, 42, 1622–1629.
- Calamini, B.; Ratia, K.; Malkowski, M.G.; Cuendet, M.; Pezzuto, J.M.; Santarsiero, B.D.; Mesecar, A.D. Pleiotropic mechanisms facilitated by resveratrol and its metabolites. Biochem. J. 2010, 429, 273–282.
- Hwang, S.H.; Wecksler, A.T.; Wagner, K.; Hammock, B.D. Rationally designed multitarget agents against inflammation and pain. Curr. Med. Chem. 2013, 20, 1783–1799.
- Lançon, A.; Frazzi, R.; Latruffe, N. Anti-oxidant, anti-inflammatory and anti-angiogenic properties of resveratrol in ocular diseases. Molecules 2016, 21, 304.
- Cheng, T.M.; Chin, Y.T.; Ho, Y.; Chen, Y.R.; Yang, Y.N.; Yang, Y.C.; Shih, Y.J.; Lin, T.I.; Lin, H.Y.; Davis, P.J. Resveratrol induces sumoylated COX-2-dependent anti-proliferation in human prostate cancer LNCaP cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 112, 67–75.
- Gong, W.H.; Zhao, N.; Zhang, Z.M.; Zhang, Y.X.; Yan, L.; Li, J.B. The inhibitory effect of resveratrol on COX-2 expression in human colorectal cancer: A promising therapeutic strategy. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1136–1143.
- Zykova, T.A.; Zhu, F.; Zhai, X.; Ma, W.Y.; Ermakova, S.P.; Lee, K.W.; Bode, A.M.; Dong, Z. Resveratrol directly targets COX-2 to inhibit carcinogenesis. Mol. Carcinog. 2008, 47, 797–805.
- Latruffe, N.; Lançon, A.; Frazzi, R.; Aires, V.; Delmas, D.; Michaille, J.J.; Djouadi, F.; Bastin, J.; Cherkaoui-Malki, M. Exploring new ways of regulation by resveratrol involving miRNAs, with emphasis on inflammation. Ann. N. Y. Acad. Sci. 2015, 1348, 97–106.
- Kim, O.Y.; Chung, J.Y.; Song, J. Effect of resveratrol on adipokines and myokines involved in fat browning: Perspectives in healthy weight against obesity. Pharm. Res. 2019, 148, 104411.
- Ye, G.; Chen, G.; Gao, H.; Lin, Y.; Liao, X.; Zhang, H.; Liu, X.; Chi, Y.; Huang, Q.; Zhu, H.; et al. Resveratrol inhibits lipid accumulation in the intestine of atherosclerotic mice and macrophages. J. Cell. Mol. Med. 2019, 23, 4313–4325.
- Feng, L.; Zhang, L. Resveratrol Suppresses Aβ-Induced Microglial Activation Through the TXNIP/TRX/NLRP3 Signaling Pathway. DNA Cell Biol. 2019, 38, 874–879.
- Ramalingam, A.; Santhanathas, T.; Shaukat Ali, S.; Zainalabidin, S. Resveratrol Supplementation Protects Against Nicotine-Induced Kidney Injury. Int. J. Environ. Res. Public Health 2019, 16, 4445.
- Chen, T.S.; Kuo, C.H.; Day, C.H.; Pan, L.F.; Chen, R.J.; Chen, B.C.; Padma, V.V.; Lin, Y.M.; Huang, C.Y. Resveratrol increases stem cell function in the treatment of damaged pancreas. J. Cell. Physiol. 2019, 234, 20443–20452.
- Wang, Y.; Wang, B.; Qi, X.; Zhang, X.; Ren, K. Resveratrol Protects Against Post-Contrast Acute Kidney Injury in Rabbits With Diabetic Nephropathy. Front. Pharmacol. 2019, 10, 833.
- Lieben Louis, X.; Raj, P.; Meikle, Z.; Yu, L.; Susser, S.E.; MacInnis, S.; Duhamel, T.A.; Wigle, J.T.; Netticadan, T. Resveratrol prevents palmitic-acid-induced cardiomyocyte contractile impairment. Can. J. Physiol. Pharmacol. 2019, 97, 1132–1140.
- Gimeno-Mallench, L.; Mas-Bargues, C.; Inglés, M.; Olaso, G.; Borras, C.; Gambini, J.; Vina, J. Resveratrol shifts energy metabolism to increase lipid oxidation in healthy old mice. Biomed. Pharmacother. 2019, 118, 109130.
- Leppäranta, O.; Sens, C.; Salmenkivi, K.; Kinnula, V.L.; Keski-Oja, J.; Myllärniemi, M.; Koli, K. Regulation of TGF-β storage and activation in the human idiopathic pulmonary fibrosis lung. Cell Tissue Res. 2012, 348, 491–503.
- Bellaye, P.S.; Yanagihara, T.; Granton, E.; Sato, S.; Shimbori, C.; Upagupta, C.; Imani, J.; Hambly, N.; Ask, K.; Gauldie, J.; et al. Macitentan reduces progression of TGF-β1-induced pulmonary fibrosis and pulmonary hypertension. Eur. Respir. J. 2018, 52.
- Wang, J.; He, F.; Chen, L.; Li, Q.; Jin, S.; Zheng, H.; Lin, J.; Zhang, H.; Ma, S.; Mei, J.; et al. Resveratrol inhibits pulmonary fibrosis by regulating miR-21 through MAPK/AP-1 pathways. Biomed. Pharmacother. 2018, 105, 37–44.
- Gao, C.; Howard-Quijano, K.; Rau, C.; Takamiya, T.; Song, Y.; Shivkumar, K.; Wang, Y.; Mahajan, A. Inflammatory and apoptotic remodeling in autonomic nervous system following myocardial infarction. PLoS ONE 2017, 12, e0177750.
- Gao, H.; Bo, Z.; Wang, Q.; Luo, L.; Zhu, H.; Ren, Y. Salvanic acid B inhibits myocardial fibrosis through regulating TGF-β1/Smad signaling pathway. Biomed. Pharmacother. 2019, 110, 685–691.
- Zhang, Y.; Lu, Y.; Ong’achwa, M.J.; Ge, L.; Qian, Y.; Chen, L.; Hu, X.; Li, F.; Wei, H.; Zhang, C.; et al. Resveratrol Inhibits the TGF-β1-Induced Proliferation of Cardiac Fibroblasts and Collagen Secretion by Downregulating miR-17 in Rat. Biomed. Res. Int. 2018, 2018, 8730593.
- Annaldas, S.; Saifi, M.A.; Khurana, A.; Godugu, C. Nimbolide ameliorates unilateral ureteral obstruction-induced renal fibrosis by inhibition of TGF-β and EMT/Slug signalling. Mol. Immunol. 2019, 112, 247–255.
- Song, M.K.; Lee, J.H.; Ryoo, I.G.; Lee, S.H.; Ku, S.K.; Kwak, M.K. Bardoxolone ameliorates TGF-β1-associated renal fibrosis through Nrf2/Smad7 elevation. Free Radic. Biol. Med. 2019, 138, 33–42.
- Zhang, X.; Lu, H.; Xie, S.; Wu, C.; Guo, Y.; Xiao, Y.; Zheng, S.; Zhu, H.; Zhang, Y.; Bai, Y. Resveratrol suppresses the myofibroblastic phenotype and fibrosis formation in kidneys via proliferation-related signalling pathways. Br. J. Pharmacol. 2019, 176, 4745–4759.
- Liu, Y. New insights into epithelial-mesenchymal transition in kidney fibrosis. J. Am. Soc. Nephrol. 2010, 21, 212–222.
- Alpers, C.E.; Hudkins, K.L.; Floege, J.; Johnson, R.J. Human renal cortical interstitial cells with some features of smooth muscle cells participate in tubulointerstitial and crescentic glomerular injury. J. Am. Soc. Nephrol. 1994, 5, 201–209.
- Bi, W.; Xu, G.; Lv, L.; Yang, C. The ratio of transforming growth factor-β1/bone morphogenetic protein-7 in the progression of the epithelial-mesenchymal transition contributes to rat liver fibrosis. Genet. Mol. Res. 2014, 13, 1005–1014.
- Kalluri, R.; Weinberg, R.A. The basics of epithelial-mesenchymal transition. J. Clin. Investig. 2009, 119, 1420–1428.
- Bai, Y.; Lu, H.; Wu, C.; Liang, Y.; Wang, S.; Lin, C.; Chen, B.; Xia, P. Resveratrol inhibits epithelial-mesenchymal transition and renal fibrosis by antagonizing the hedgehog signaling pathway. Biochem. Pharmacol. 2014, 92, 484–493.
- Zhai, X.X.; Ding, J.C.; Tang, Z.M. Resveratrol Inhibits Proliferation and Induces Apoptosis of Pathological Scar Fibroblasts Through the Mechanism Involving TGF-β1/Smads Signaling Pathway. Cell Biochem. Biophys. 2015, 71, 1267–1272.
- Liu, S.; Zhao, M.; Zhou, Y.; Wang, C.; Yuan, Y.; Li, L.; Bresette, W.; Chen, Y.; Cheng, J.; Lu, Y.; et al. Resveratrol exerts dose-dependent anti-fibrotic or pro-fibrotic effects in kidneys: A potential risk to individuals with impaired kidney function. Phytomedicine 2019, 57, 223–235.
- Chávez, E.; Reyes-Gordillo, K.; Segovia, J.; Shibayama, M.; Tsutsumi, V.; Vergara, P.; Moreno, M.G.; Muriel, P. Resveratrol prevents fibrosis, NF-kappaB activation and TGF-beta increases induced by chronic CCl4 treatment in rats. J. Appl. Toxicol. JAT 2008, 28, 35–43.
- Ding, S.; Wang, H.; Wang, M.; Bai, L.; Yu, P.; Wu, W. Resveratrol alleviates chronic “real-world” ambient particulate matter-induced lung inflammation and fibrosis by inhibiting NLRP3 inflammasome activation in mice. Ecotoxicol. Environ. Saf. 2019, 182, 109425.
- Sun, D.Y.; Wu, J.Q.; He, Z.H.; He, M.F.; Sun, H.B. Cancer-associated fibroblast regulate proliferation and migration of prostate cancer cells through TGF-β signaling pathway. Life Sci. 2019, 235, 116791.
- Cruz-Bermúdez, A.; Laza-Briviesca, R.; Vicente-Blanco, R.J.; García-Grande, A.; Coronado, M.J.; Laine-Menéndez, S.; Alfaro, C.; Sanchez, J.C.; Franco, F.; Calvo, V.; et al. Cancer-associated fibroblasts modify lung cancer metabolism involving ROS and TGF-β signaling. Free Radic. Biol. Med. 2019, 130, 163–173.
- Bierie, B.; Moses, H.L. TGF-beta and cancer. Cytokine Growth Factor Rev. 2006, 17, 29–40.
- Zhao, M.; Mishra, L.; Deng, C.X. The role of TGF-β/SMAD4 signaling in cancer. Int. J. Biol. Sci. 2018, 14, 111–123.
- Camerlingo, R.; Miceli, R.; Marra, L.; Rea, G.; D’Agnano, I.; Nardella, M.; Montella, R.; Morabito, A.; Normanno, N.; Tirino, V.; et al. Conditioned medium of primary lung cancer cells induces EMT in A549 lung cancer cell line by TGF-ß1 and miRNA21 cooperation. PLoS ONE 2019, 14, e0219597.
- Liu, X.S.; Lin, X.K.; Mei, Y.; Ahmad, S.; Yan, C.X.; Jin, H.L.; Yu, H.; Chen, C.; Lin, C.Z.; Yu, J.R. Regulatory T Cells Promote Overexpression of Lgr5 on Gastric Cancer Cells via TGF-beta1 and Confer Poor Prognosis in Gastric Cancer. Front. Immunol. 2019, 10, 1741.
- Hyun Lee, J.; Dhananjaya Mohan, C.; Deivasigamani, A.; Yun Jung, Y.; Rangappa, S.; Basappa, S.; Chinnathambi, A.; Awad Alahmadi, T.; Ali Alharbi, S.; Garg, M.; et al. Brusatol suppresses STAT3-driven metastasis by downregulating epithelial-mesenchymal transition in hepatocellular carcinoma. J. Adv. Res. 2020.
- Cheng, J.-T.; Wang, L.; Wang, H.; Tang, F.-R.; Cai, W.-Q.; Sethi, G.; Xin, H.-W.; Ma, Z. Insights into Biological Role of LncRNAs in Epithelial-Mesenchymal Transition. Cells 2019, 8, 1178.
- Loh, C.-Y.; Chai, J.Y.; Tang, T.F.; Wong, W.F.; Sethi, G.; Shanmugam, M.K.; Chong, P.P.; Looi, C.Y. The E-Cadherin and N-Cadherin Switch in Epithelial-to-Mesenchymal Transition: Signaling, Therapeutic Implications, and Challenges. Cells 2019, 8, 1118.
- Sun, Y.; Zhou, Q.M.; Lu, Y.Y.; Zhang, H.; Chen, Q.L.; Zhao, M.; Su, S.B. Resveratrol Inhibits the Migration and Metastasis of MDA-MB-231 Human Breast Cancer by Reversing TGF-β1-Induced Epithelial-Mesenchymal Transition. Molecules 2019, 24, 1131.
- Wang, H.; Zhang, H.; Tang, L.; Chen, H.; Wu, C.; Zhao, M.; Yang, Y.; Chen, X.; Liu, G. Resveratrol inhibits TGF-β1-induced epithelial-to-mesenchymal transition and suppresses lung cancer invasion and metastasis. Toxicology 2013, 303, 139–146.
- Song, Y.; Chen, Y.; Li, Y.; Lyu, X.; Cui, J.; Cheng, Y.; Zheng, T.; Zhao, L.; Zhao, G. Resveratrol Suppresses Epithelial-Mesenchymal Transition in GBM by Regulating Smad-Dependent Signaling. Biomed. Res. Int. 2019, 2019, 1321973.
- Kabel, A.M.; Atef, A.; Estfanous, R.S. Ameliorative potential of sitagliptin and/or resveratrol on experimentally-induced clear cell renal cell carcinoma. Biomed. Pharmacother. 2018, 97, 667–674.
- Zhang, Y.; Yang, S.; Yang, Y.; Liu, T. Resveratrol induces immunogenic cell death of human and murine ovarian carcinoma cells. Infect. Agents Cancer 2019, 14, 27.
- Rekik, R.; Belhadj Hmida, N.; Ben Hmid, A.; Zamali, I.; Kammoun, N.; Ben Ahmed, M. PD-1 induction through TCR activation is partially regulated by endogenous TGF-β. Cell. Mol. Immunol. 2015, 12, 648–649.
- Celada, L.J.; Kropski, J.A.; Herazo-Maya, J.D.; Luo, W.; Creecy, A.; Abad, A.T.; Chioma, O.S.; Lee, G.; Hassell, N.E.; Shaginurova, G.I.; et al. PD-1 up-regulation on CD4(+) T cells promotes pulmonary fibrosis through STAT3-mediated IL-17A and TGF-β1 production. Sci. Transl. Med. 2018, 10.
- Johnson, E.R.; Matthay, M.A. Acute lung injury: Epidemiology, pathogenesis, and treatment. J. Aerosol Med. Pulm. Drug Deliv. 2010, 23, 243–252.
- Sawa, T. The molecular mechanism of acute lung injury caused by Pseudomonas aeruginosa: From bacterial pathogenesis to host response. J. Intensive Care 2014, 2, 10.
- Kubota, Y.; Iwasaki, Y.; Harada, H.; Yokomura, I.; Ueda, M.; Hashimoto, S.; Nakagawa, M. Role of alveolar macrophages in Candida-induced acute lung injury. Clin. Diagn. Lab. Immunol. 2001, 8, 1258–1262.
- Savransky, V.; Rostapshov, V.; Pinelis, D.; Polotsky, Y.; Korolev, S.; Komisar, J.; Fegeding, K. Murine lethal toxic shock caused by intranasal administration of staphylococcal enterotoxin B. Toxicol. Pathol. 2003, 31, 373–378.
- Alghetaa, H.; Mohammed, A.; Sultan, M.; Busbee, P.; Murphy, A.; Chatterjee, S.; Nagarkatti, M.; Nagarkatti, P. Resveratrol protects mice against SEB-induced acute lung injury and mortality by miR-193a modulation that targets TGF-β signalling. J. Cell. Mol. Med. 2018, 22, 2644–2655.
- Karagiannidis, C.; Akdis, M.; Holopainen, P.; Woolley, N.J.; Hense, G.; Rückert, B.; Mantel, P.Y.; Menz, G.; Akdis, C.A.; Blaser, K.; et al. Glucocorticoids upregulate FOXP3 expression and regulatory T cells in asthma. J. Allergy Clin. Immunol. 2004, 114, 1425–1433.
- Alharris, E.; Alghetaa, H.; Seth, R.; Chatterjee, S.; Singh, N.P.; Nagarkatti, M.; Nagarkatti, P. Resveratrol Attenuates Allergic Asthma and Associated Inflammation in the Lungs Through Regulation of miRNA-34a That Targets FoxP3 in Mice. Front. Immunol. 2018, 9, 2992.
- Wollin, L.; Pieper, M. Tiotropium bromide exerts anti-inflammatory activity in a cigarette smoke mouse model of COPD. Pulm. Pharmacol. Ther. 2010, 23, 345–354.
- Tamimi, A.; Serdarevic, D.; Hanania, N.A. The effects of cigarette smoke on airway inflammation in asthma and COPD: Therapeutic implications. Respir. Med. 2012, 106, 319–328.
- Busse, P.J.; Zhang, T.F.; Srivastava, K.; Lin, B.P.; Schofield, B.; Sealfon, S.C.; Li, X.-M. Chronic exposure to TNF-α increases airway mucus gene expression in vivo. J. Allergy Clin. Immunol. 2005, 116, 1256–1263.
- Numasaki, M.; Tomioka, Y.; Takahashi, H.; Sasaki, H. IL-17 and IL-17F modulate GM-CSF production by lung microvascular endothelial cells stimulated with IL-1β and/or TNF-α. Immunol. Lett. 2004, 95, 175–184.
- Chen, J.; Yang, X.; Zhang, W.; Peng, D.; Xia, Y.; Lu, Y.; Han, X.; Song, G.; Zhu, J.; Liu, R. Therapeutic Effects of Resveratrol in a Mouse Model of LPS and Cigarette Smoke-Induced COPD. Inflammation 2016, 39, 1949–1959.
- Al-Mufti, F.; Amuluru, K.; Changa, A.; Lander, M.; Patel, N.; Wajswol, E.; Al-Marsoummi, S.; Alzubaidi, B.; Singh, I.P.; Nuoman, R.; et al. Traumatic brain injury and intracranial hemorrhage-induced cerebral vasospasm: A systematic review. Neurosurg. Focus 2017, 43, E14.
- Shoamanesh, A.; Kwok, C.S.; Lim, P.A.; Benavente, O.R. Postthrombolysis intracranial hemorrhage risk of cerebral microbleeds in acute stroke patients: A systematic review and meta-analysis. Int. J. Stroke Off. J. Int. Stroke Soc. 2013, 8, 348–356.
- Bugeme, M.; Mukuku, O. Neuropsychiatric manifestations revealing cerebral subarachnoid hemorrhage caused by electrification accident about a case and review of literature. Pan Afr. Med. J. 2014, 18, 201.
- Logan, T.T.; Villapol, S.; Symes, A.J. TGF-β superfamily gene expression and induction of the Runx1 transcription factor in adult neurogenic regions after brain injury. PLoS ONE 2013, 8, e59250.
- Zhao, R.; Zhao, K.; Su, H.; Zhang, P.; Zhao, N. Resveratrol ameliorates brain injury via the TGF-β-mediated ERK signaling pathway in a rat model of cerebral hemorrhage. Exp. Ther. Med. 2019, 18, 3397–3404.
- Tiwari, V.; Chopra, K. Resveratrol prevents alcohol-induced cognitive deficits and brain damage by blocking inflammatory signaling and cell death cascade in neonatal rat brain. J. Neurochem. 2011, 117, 678–690.
- Forbes, J.M.; Cooper, M.E.; Oldfield, M.D.; Thomas, M.C. Role of advanced glycation end products in diabetic nephropathy. J. Am. Soc. Nephrol. 2003, 14, S254–S258.
- Yamagishi, S.-i.; Matsui, T. Advanced glycation end products, oxidative stress and diabetic nephropathy. Oxidative Med. Cell. Longev. 2010, 3, 101–108.
- Mima, A. Inflammation and oxidative stress in diabetic nephropathy: New insights on its inhibition as new therapeutic targets. J. Diabetes Res. 2013, 2013, 8.
- Hussein, M.M.; Mahfouz, M.K. Effect of resveratrol and rosuvastatin on experimental diabetic nephropathy in rats. Biomed. Pharmacother. 2016, 82, 685–692.
- Chen, K.H.; Hung, C.C.; Hsu, H.H.; Jing, Y.H.; Yang, C.W.; Chen, J.K. Resveratrol ameliorates early diabetic nephropathy associated with suppression of augmented TGF-β/smad and ERK1/2 signaling in streptozotocin-induced diabetic rats. Chem. Biol. Interact. 2011, 190, 45–53.
More
Information
Subjects:
Medicine, General & Internal
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
06 Jul 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No