 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Lijun Ding | + 1916 word(s) | 1916 | 2021-06-03 09:41:42 | | | |
| 2 | Enzi Gong | Meta information modification | 1916 | 2021-06-07 11:18:09 | | | | |
| 3 | Enzi Gong | Meta information modification | 1916 | 2021-06-07 11:21:32 | | | | |
| 4 | Enzi Gong | Meta information modification | 1916 | 2021-06-07 11:26:01 | | |
Video Upload Options
Ovarian steroid-regulated cyclical regeneration of the endometrium is crucial for endometrial receptivity and embryo implantation, and it is dependent on the dynamic remodeling of the endometrial vasculature. Perivascular cells, including pericytes surrounding capillaries and microvessels and adventitial cells located in the outermost layer of large vessels, show properties of mesenchymal stem cells, and they are thus promising candidates for uterine regeneration.
1. Overview
Ovarian steroid-regulated cyclical regeneration of the endometrium is crucial for endometrial receptivity and embryo implantation, and it is dependent on the dynamic remodeling of the endometrial vasculature. Perivascular cells, including pericytes surrounding capillaries and microvessels and adventitial cells located in the outermost layer of large vessels, show properties of mesenchymal stem cells, and they are thus promising candidates for uterine regeneration. In this review, we discuss the structure and functions of the endometrial blood vasculature and their roles in endometrial regeneration, the main biomarkers and characteristics of perivascular cells in the endometrium, and stem cell-based angiogenetic therapy for Asherman’s syndrome.
2. Introduction
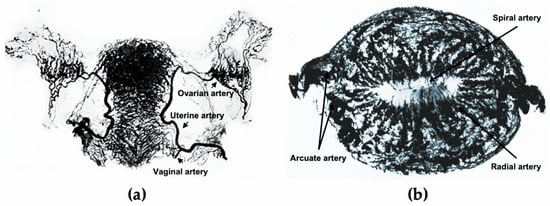
3. The Endometrial Vasculature
4. Conclusions
In conclusion, the progenitor role of perivascular cells derived from human endometrium and a stem cell-based therapy for AS have been investigated in both murine model studies and clinical trials. Nevertheless, several crucial questions have yet to be solved. First, the relationship between human endometrial vasculature and uterine development under physiological conditions remains unknown. The synchronous development of uterus and blood vessels is to be further revealed by combining the means of embryo anatomy, multi-omics analysis, and single-cell sequencing. Second, the role of perivascular cells during the processes of uterine development and cyclic endometrial regeneration needs to be further explored by in vivo cell lineage tracing. Third, by combining with tissue engineering, perivascular cells are promising candidates for tissue repair under pathological conditions, while better seed cells and improved delivery methods need to be explored. Moreover, the mechanisms behind how the precursor cells mobilize the remaining local cells and how they induce microenvironmental changes to allow themselves to colonize remain unclear.
References
- Maybin, J.A.; Critchley, H.O.D. Menstrual Physiology: Implications for Endometrial Pathology and Beyond. Hum. Reprod. Update 2015, 21, 748–761.
- Mihm, M.; Gangooly, S.; Muttukrishna, S. The Normal Menstrual Cycle in Women. Anim. Reprod. Sci. 2011, 124, 229–236.
- Samimi, M.; Pourhanifeh, M.H.; Mehdizadehkashi, A.; Eftekhar, T.; Asemi, Z. The Role of Inflammation, Oxidative Stress, Angiogenesis, and Apoptosis in the Pathophysiology of Endometriosis: Basic Science and New Insights Based on Gene Expression. J. Cell Physiol. 2019, 234, 19384–19392.
- Buzzaccarini, G.; Vitagliano, A.; Andrisani, A.; Santarsiero, C.M.; Cicinelli, R.; Nardelli, C.; Ambrosini, G.; Cicinelli, E. Chronic Endometritis and Altered Embryo Implantation: A Unified Pathophysiological Theory from a Literature Systematic Review. J. Assist. Reprod Genet. 2020, 37, 2897–2911.
- Gargett, C.E.; Chan, R.W.S.; Schwab, K.E. Endometrial Stem Cells. Curr. Opin. Obstet. Gynecol. 2007, 19, 377–383.
- Friedler, S.; Margalioth, E.J.; Kafka, I.; Yaffe, H. Incidence of Post-Abortion Intra-Uterine Adhesions Evaluated by Hysteroscopy---A Prospective Study. Hum. Reprod 1993, 8, 442–444.
- Badawy, S.Z.; Orbuch, L.; Khurana, K.K. Secondary Amenorrhea with Severe Intrauterine Adhesions and Chronic Uterine Torsion after Cesarean Section in a Teenage Girl. J. Pediatr. Adolesc. Gynecol. 1998, 11, 93–96.
- Schenker, J.G.; Margalioth, E.J. Intrauterine Adhesions: An Updated Appraisal. Fertil. Steril. 1982, 37, 593–610.
- Rabau, E.; David, A. Intrauterine adhesions: Etiology, prevention, and treatment. Obstet. Gynecol. 1963, 22, 626–629.
- Deane, J.A.; Gualano, R.C.; Gargett, C.E. Regenerating Endometrium from Stem/Progenitor Cells: Is It Abnormal in Endometriosis, Asherman’s Syndrome and Infertility? Curr. Opin. Obstet. Gynecol. 2013, 25, 193–200.
- Arey, L.B. Developmental Anatomy: A Textbook and Laboratory Manual of Embryology; W. B. Saunders Company: Philadelphia, PA, USA, 1974; ISBN 978-0-7216-3443-2.
- Farrer-Brown, G.; Beilby, J.O.; Tarbit, M.H. The Blood Supply of the Uterus. 1. Arterial Vasculature. J. Obstet. Gynaecol. Br. Commonw. 1970, 77, 673–681.
- Herbert, S.P.; Stainier, D.Y.R. Molecular Control of Endothelial Cell Behaviour during Blood Vessel Morphogenesis. Nat. Rev. Mol. Cell Biol. 2011, 12, 551–564.
- Potente, M.; Mäkinen, T. Vascular Heterogeneity and Specialization in Development and Disease. Nat. Rev. Mol. Cell Biol. 2017, 18, 477–494.
- Stratman, A.N.; Yu, J.A.; Mulligan, T.S.; Butler, M.G.; Sause, E.T.; Weinstein, B.M. Chapter 24—Blood Vessel Formation. In Principles of Developmental Genetics, 2nd ed.; Moody, S.A., Ed.; Academic Press: Oxford, UK, 2015; pp. 421–449. ISBN 978-0-12-405945-0.
- Folkman, J. Angiogenesis in Cancer, Vascular, Rheumatoid and Other Disease. Nat. Med. 1995, 1, 27–31.
- Flamme, I.; Frölich, T.; Risau, W. Molecular Mechanisms of Vasculogenesis and Embryonic Angiogenesis. J. Cell Physiol. 1997, 173, 206–210.
- Bikfalvi, A. History and Conceptual Developments in Vascular Biology and Angiogenesis Research: A Personal View. Angiogenesis 2017, 20, 463–478.
- Cicinelli, E.; Einer-Jensen, N.; Galantino, P.; Alfonso, R.; Nicoletti, R. The Vascular Cast of the Human Uterus: From Anatomy to Physiology. Ann. N. Y. Acad. Sci. 2004, 1034, 19–26.
- Smith, S.K. Regulation of Angiogenesis in the Endometrium. Trends Endocrinol. Metab. 2001, 12, 147–151.
- Gargett, C.E.; Rogers, P.A. Human Endometrial Angiogenesis. Reproduction 2001, 121, 181–186.
- Risau, W. Mechanisms of Angiogenesis. Nature 1997, 386, 671–674.
- Asahara, T.; Masuda, H.; Takahashi, T.; Kalka, C.; Pastore, C.; Silver, M.; Kearne, M.; Magner, M.; Isner, J.M. Bone Marrow Origin of Endothelial Progenitor Cells Responsible for Postnatal Vasculogenesis in Physiological and Pathological Neovascularization. Circ Res. 1999, 85, 221–228.
- Burri, P.H.; Djonov, V. Intussusceptive Angiogenesis--the Alternative to Capillary Sprouting. Mol. Asp. Med. 2002, 23, S1–S27.
- Gambino, L.S.; Wreford, N.G.; Bertram, J.F.; Dockery, P.; Lederman, F.; Rogers, P.A.W. Angiogenesis Occurs by Vessel Elongation in Proliferative Phase Human Endometrium. Hum. Reprod. 2002, 17, 1199–1206.
- Goodger, A.M.; Rogers, P.A. Endometrial Endothelial Cell Proliferation during the Menstrual Cycle. Hum. Reprod. 1994, 9, 399–405.
- Ferenczy, A.; Bertrand, G.; Gelfand, M.M. Proliferation Kinetics of Human Endometrium during the Normal Menstrual Cycle. Am. J. Obstet. Gynecol. 1979, 133, 859–867.
- Rogers, P.A.; Lederman, F.; Taylor, N. Endometrial Microvascular Growth in Normal and Dysfunctional States. Hum. Reprod. Update 1998, 4, 503–508.
- Nayak, N.R.; Brenner, R.M. Vascular Proliferation and Vascular Endothelial Growth Factor Expression in the Rhesus Macaque Endometrium. J. Clin. Endocrinol. Metab. 2002, 87, 1845–1855.
- Limbourg, A.; Korff, T.; Napp, L.C.; Schaper, W.; Drexler, H.; Limbourg, F.P. Evaluation of Postnatal Arteriogenesis and Angiogenesis in a Mouse Model of Hind-Limb Ischemia. Nat. Protoc. 2009, 4, 1737–1746.
- Girling, J.E.; Lederman, F.L.; Walter, L.M.; Rogers, P.A.W. Progesterone, but Not Estrogen, Stimulates Vessel Maturation in the Mouse Endometrium. Endocrinology 2007, 148, 5433–5441.
- Jabbour, H.N.; Kelly, R.W.; Fraser, H.M.; Critchley, H.O.D. Endocrine Regulation of Menstruation. Endocr. Rev. 2006, 27, 17–46.
- Kooy, J.; Taylor, N.H.; Healy, D.L.; Rogers, P.A.W. Endothelial Cell Proliferation in the Endometrium of Women with Menorrhagia and in Women Following Endometrial Ablation. Hum. Reprod. 1996, 11, 1067–1072.
- Nisolle, M.; Casanas-Roux, F.; Anaf, V.; Mine, J.M.; Donnez, J. Morphometric Study of the Stromal Vascularization in Peritoneal Endometriosis. Fertil. Steril. 1993, 59, 681–684.
- Oosterlynck, D.J.; Meuleman, C.; Sobis, H.; Vandeputte, M.; Koninckx, P.R. Angiogenic Activity of Peritoneal Fluid from Women with Endometriosis. Fertil. Steril. 1993, 59, 778–782.
- Jiang, P.; Tang, X.; Wang, H.; Dai, C.; Su, J.; Zhu, H.; Song, M.; Liu, J.; Nan, Z.; Ru, T.; et al. Collagen-Binding Basic Fibroblast Growth Factor Improves Functional Remodeling of Scarred Endometrium in Uterine Infertile Women: A Pilot Study. Sci. China Life Sci. 2019, 62, 1617–1629.

