 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Chongshan Dai | + 2027 word(s) | 2027 | 2021-05-28 05:25:32 | | | |
| 2 | Vivi Li | Meta information modification | 2027 | 2021-05-28 07:39:51 | | |
Video Upload Options
The emergence of antimicrobial resistance in Gram-negative bacteria poses a huge health challenge. The therapeutic use of polymyxins (i.e., colistin and polymyxin B) is commonplace due to high efficacy and limiting treatment options for multidrug-resistant Gram-negative bacterial infections. Nephrotoxicity and neurotoxicity are the major dose-limiting factors that limit the therapeutic window of polymyxins; nephrotoxicity is a complication in up to ~60% of patients. The emergence of polymyxin-resistant strains or polymyxin heteroresistance is also a limiting factor. These caveats have catalyzed the search for polymyxin combinations that synergistically kill polymyxin-susceptible and resistant organisms and/or minimize the unwanted side effects. Curcumin—an FDA-approved natural product—exerts many pharmacological activities. Recent studies showed that polymyxins–curcumin combinations showed a synergistically inhibitory effect on the growth of bacteria (e.g., Gram-positive and Gram-negative bacteria) in vitro. Moreover, curcumin co-administration ameliorated colistin-induced nephrotoxicity and neurotoxicity by inhibiting oxidative stress, mitochondrial dysfunction, inflammation and apoptosis.
1. Introduction
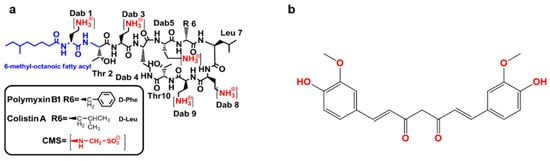
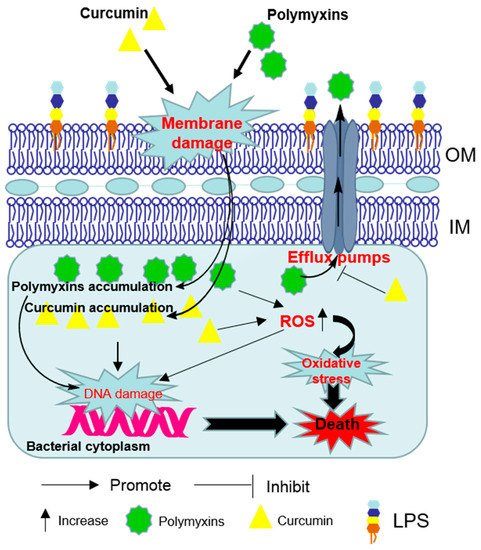
2. Synergistic Antibacterial Effects of the Polymyxin in Combination with Curcumin
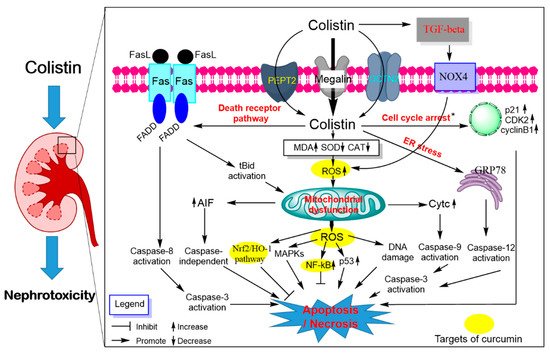
3. Polymyxin-Induced Nephrotoxicity and Protective Effect of Curcumin
References
- The World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; WHO: Geneva, Switzerland, 2017.
- Tyers, M.; Wright, G.D. Drug combinations: A strategy to extend the life of antibiotics in the 21st century. Nat. Rev. Microbiol. 2019, 17, 141–155.
- Koyama, Y.; Kurosasa, A.; Tsuchiya, A.; Takakuta, K. A new antibiotic “colistin” produced by spore-forming soil bacteria. J. Antibiot. 1950, 3, 457–458.
- Braine, T. Race against time to develop new antibiotics. Bull. World Health Organ. 2011, 89, 88–89.
- Zowawi, H.M.; Harris, P.N.; Roberts, M.J.; Tambyah, P.A.; Schembri, M.A.; Pezzani, M.D.; Williamson, D.A.; Paterson, D.L. The emerging threat of multidrug-resistant gram-negative bacteria in urology. Nat. Rev. Urol. 2015, 12, 570–584.
- Demiraslan, H.; Cevahir, F.; Berk, E.; Metan, G.; Cetin, M.; Alp, E. Is surveillance for colonization of carbapenem-resistant gram-negative bacteria important in adult bone marrow transplantation units? Am. J. Infect. Control 2017, 45, 735–739.
- Karlowsky, J.A.; Lob, S.H.; Kazmierczak, K.M.; Hawser, S.P.; Magnet, S.; Young, K.; Motyl, M.R.; Sahm, D.F. In vitro activity of imipenem/relebactam against gram-negative eskape pathogens isolated in 17 european countries: 2015 smart surveillance programme. J. Antimicrob. Chemother. 2018, 73, 1872–1879.
- Adams-Sapper, S.; Gayoso, A.; Riley, L.W. Stress-adaptive responses associated with high-level carbapenem resistance in kpc-producing klebsiella pneumoniae. J. Pathog. 2018, 2018, 11.
- Exner, M.; Bhattacharya, S.; Christiansen, B.; Gebel, J.; Goroncy-Bermes, P.; Hartemann, P.; Heeg, P.; Ilschner, C.; Kramer, A.; Larson, E.; et al. Antibiotic resistance: What is so special about multidrug-resistant gram-negative bacteria? GMS Hyg. Infect. Control 2017, 12, Doc05.
- Oliveira, V.D.; Rubio, F.G.; Almeida, M.T.; Nogueira, M.C.; Pignatari, A.C. Trends of 9416 multidrug-resistant gram-negative bacteria. Rev. Assoc. Med. Bras. 2015, 61, 244–249.
- Perez, F.; Adachi, J.; Bonomo, R.A. Antibiotic-resistant gram-negative bacterial infections in patients with cancer. Clin. Infect. Dis. 2014, 59, S335–S339.
- Nation, R.L.; Velkov, T.; Li, J. Colistin and polymyxin b: Peas in a pod, or chalk and cheese? Clin. Infect. Dis. 2014, 59, 88–94.
- Nation, R.L.; Garonzik, S.M.; Thamlikitkul, V.; Giamarellos-Bourboulis, E.J.; Forrest, A.; Paterson, D.L.; Li, J.; Silveira, F.P. Dosing guidance for intravenous colistin in critically-ill patients. Clin. Infect. Dis. 2017, 64, 565–571.
- Li, J.; Nation, R.L.; Turnidge, J.D.; Milne, R.W.; Coulthard, K.; Rayner, C.R.; Paterson, D.L. Colistin: The re-emerging antibiotic for multidrug-resistant gram-negative bacterial infections. Lancet Infect. Dis. 2006, 6, 589–601.
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism mcr-1 in animals and human beings in china: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168.
- Napier, B.A.; Band, V.; Burd, E.M.; Weiss, D.S. Colistin heteroresistance in enterobacter cloacae is associated with cross-resistance to the host antimicrobial lysozyme. Antimicrob. Agents Chemother. 2014, 58, 5594–5597.
- Garonzik, S.M.; Li, J.; Thamlikitkul, V.; Paterson, D.L.; Shoham, S.; Jacob, J.; Silveira, F.P.; Forrest, A.; Nation, R.L. Population pharmacokinetics of colistin methanesulfonate and formed colistin in critically ill patients from a multicenter study provide dosing suggestions for various categories of patients. Antimicrob. Agents Chemother. 2011, 55, 3284–3294.
- Hartzell, J.D.; Neff, R.; Ake, J.; Howard, R.; Olson, S.; Paolino, K.; Vishnepolsky, M.; Weintrob, A.; Wortmann, G. Nephrotoxicity associated with intravenous colistin (colistimethate sodium) treatment at a tertiary care medical center. Clin. Infect. Dis. 2009, 48, 1724–1728.
- Ni, W.T.; Shao, X.D.; Di, X.Z.; Cui, J.C.; Wang, R.; Liu, Y.N. In vitro synergy of polymyxins with other antibiotics for acinetobacter Baumannii: A systematic review and meta-analysis. Int. J. Antimicrob. Agents 2015, 45, 8–18.
- O’Hara, J.A.; Ambe, L.A.; Casella, L.G.; Townsend, B.M.; Pelletier, M.R.; Ernst, R.K.; Shanks, R.M.Q.; Doi, Y. Activities of vancomycin-containing regimens against colistin-resistant acinetobacter Baumannii clinical strains. Antimicrob. Agents Chemother. 2013, 57, 2103–2108.
- Salam, A.M.; Quave, C.L. Opportunities for plant natural products in infection control. Curr. Opin. Microbiol. 2018, 45, 189–194.
- Silver, L.L. Natural products as a source of drug leads to overcome drug resistance. Future Microbiol. 2015, 10, 1711–1718.
- Betts, J.W.; Sharili, A.S.; La Ragione, R.M.; Wareham, D.W. In vitro antibacterial activity of curcumin-polymyxin b combinations against multidrug-resistant bacteria associated with traumatic wound infections. J. Nat. Prod. 2016, 79, 1702–1706.
- Kaur, A.; Sharma, P.; Capalash, N. Curcumin alleviates persistence of acinetobacter Baumannii against colistin. Sci. Rep. 2018, 8, 11029.
- Dai, C.; Xiao, X.; Zhang, Y.; Xiang, B.; Hoyer, D.; Shen, J.; Velkov, T.; Tang, S. Curcumin attenuates colistin-induced peripheral neurotoxicity in mice. ACS Infect. Dis. 2020, 6, 715–724.
- Dai, C.; Ciccotosto, G.D.; Cappai, R.; Tang, S.; Li, D.; Xie, S.; Xiao, X.; Velkov, T. Curcumin attenuates colistin-induced neurotoxicity in n2a cells via anti-inflammatory activity, suppression of oxidative stress, and apoptosis. Mol. Neurobiol. 2018, 55, 421–434.
- Edrees, N.E.; Galal, A.A.A.; Monaem, A.R.A.; Beheiry, R.R.; Metwally, M.M.M. Curcumin alleviates colistin-induced nephrotoxicity and neurotoxicity in rats via attenuation of oxidative stress, inflammation and apoptosis. Chem. Biol. Interact. 2018, 294, 56–64.
- Lin, X.P.; Xue, C.; Zhang, J.M.; Wu, W.J.; Chen, X.Y.; Zeng, Y.M. Curcumin inhibits lipopolysaccharide-induced mucin 5ac hypersecretion and airway inflammation via nuclear factor erythroid 2-related factor 2. Chin. Med. J. 2018, 131, 1686–1693.
- Lobo de Sa, F.D.; Butkevych, E.; Nattramilarasu, P.K.; Fromm, A.; Mousavi, S.; Moos, V.; Golz, J.C.; Stingl, K.; Kittler, S.; Seinige, D.; et al. Curcumin mitigates immune-induced epithelial barrier dysfunction by campylobacter Jejuni. Int. J. Mol. Sci. 2019, 20, 4830.
- Santos, A.M.; Lopes, T.; Oleastro, M.; Gato, I.V.; Floch, P.; Benejat, L.; Chaves, P.; Pereira, T.; Seixas, E.; Machado, J.; et al. Curcumin inhibits gastric inflammation induced by helicobacter pylori infection in a mouse model. Nutrients 2015, 7, 306–320.
- Schaefers, M.M.; Breshears, L.M.; Anderson, M.J.; Lin, Y.C.; Grill, A.E.; Panyam, J.; Southern, P.J.; Schlievert, P.M.; Peterson, M.L. Epithelial proinflammatory response and curcumin-mediated protection from Staphylococcal toxic shock syndrome toxin-1. PLoS ONE 2012, 7, e32813.
- Zhou, J.; Miao, H.; Li, X.; Hu, Y.; Sun, H.; Hou, Y. Curcumin inhibits placental inflammation to ameliorate lps-induced adverse pregnancy outcomes in mice via upregulation of phosphorylated akt. Inflamm. Res. 2017, 66, 177–185.
- Kumari, A.; Dash, D.; Singh, R. Curcumin inhibits Lipopolysaccharide (lps)-induced endotoxemia and airway inflammation through modulation of sequential release of inflammatory mediators (tnf-alpha and tgf-beta1) in murine model. Inflammopharmacology 2017, 25, 329–341.
- Sharma, R.A.; McLelland, H.R.; Hill, K.A.; Ireson, C.R.; Euden, S.A.; Manson, M.M.; Pirmohamed, M.; Marnett, L.J.; Gescher, A.J.; Steward, W.P. Pharmacodynamic and pharmacokinetic study of oral curcuma extract in patients with colorectal cancer. Clin. Cancer Res. 2001, 7, 1894–1900.
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818.
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic roles of curcumin: Lessons learned from clinical trials. AAPS J. 2013, 15, 195–218.
- Cheng, A.L.; Hsu, C.H.; Lin, J.K.; Hsu, M.M.; Ho, Y.F.; Shen, T.S.; Ko, J.Y.; Lin, J.T.; Lin, B.R.; Ming-Shiang, W.; et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res. 2001, 21, 2895–2900.
- Lao, C.D.; Ruffin, M.T.T.; Normolle, D.; Heath, D.D.; Murray, S.I.; Bailey, J.M.; Boggs, M.E.; Crowell, J.; Rock, C.L.; Brenner, D.E. Dose escalation of a curcuminoid formulation. BMC Complement. Altern. Med. 2006, 6, 10.
- Poirel, L.; Jayol, A.; Nordmann, P. Polymyxins: Antibacterial activity, susceptibility testing, and resistance mechanisms encoded by plasmids or chromosomes. Clin. Microbiol. Rev. 2017, 30, 557–596.
- Sampson, T.R.; Liu, X.; Schroeder, M.R.; Kraft, C.S.; Burd, E.M.; Weiss, D.S. Rapid killing of acinetobacter Baumannii by polymyxins is mediated by a hydroxyl radical death pathway. Antimicrob. Agents Chemother. 2012, 56, 5642–5649.
- Yu, Z.; Qin, W.; Lin, J.; Fang, S.; Qiu, J. Antibacterial mechanisms of polymyxin and bacterial resistance. Biomed. Res. Int. 2015, 2015, 679109.
- Needham, B.D.; Trent, M.S. Fortifying the barrier: The impact of lipid a remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 2013, 11, 467–481.
- Olaitan, A.O.; Morand, S.; Rolain, J.M. Mechanisms of polymyxin resistance: Acquired and intrinsic resistance in bacteria. Front. Microbiol. 2014, 5, 643.
- Kofteridis, D.P.; Alexopoulou, C.; Valachis, A.; Maraki, S.; Dimopoulou, D.; Georgopoulos, D.; Samonis, G. Aerosolized plus intravenous colistin versus intravenous colistin alone for the treatment of ventilator-associated pneumonia: A matched case-control study. Clin. Infect. Dis. 2010, 51, 1238–1244.
- Gai, Z.B.; Samodelov, S.L.; Kullak-Ublick, G.A.; Visentin, M. Molecular mechanisms of colistin-induced nephrotoxicity. Molecules 2019, 24, 653.
- Dezoti-Fonseca, C.; Watanabe, M.; Vattimo-Mde, F. Role of heme oxygenase-1 in polymyxin b-induced nephrotoxicity in rats. Antimicrob. Agents Chemother. 2012, 56, 5082–5087.
- Dai, C.; Tang, S.; Wang, Y.; Velkov, T.; Xiao, X. Baicalein acts as a nephroprotectant that ameliorates colistin-induced nephrotoxicity by activating the antioxidant defence mechanism of the kidneys and down-regulating the inflammatory response. J. Antimicrob. Chemother. 2017, 72, 2562–2569.
- Wallace, S.J.; Li, J.; Nation, R.L.; Rayner, C.R.; Taylor, D.; Middleton, D.; Milne, R.W.; Coulthard, K.; Turnidge, J.D. Subacute toxicity of colistin methanesulfonate in rats: Comparison of various intravenous dosage regimens. Antimicrob. Agents Chemother. 2008, 52, 1159–1161.
- Ghlissi, Z.; Hakim, A.; Mnif, H.; Ayadi, F.M.; Zeghal, K.; Rebai, T.; Sahnoun, Z. Evaluation of colistin nephrotoxicity administered at different doses in the rat model. Ren. Fail. 2013, 35, 1130–1135.
- Keirstead, N.D.; Wagoner, M.P.; Bentley, P.; Blais, M.; Brown, C.; Cheatham, L.; Ciaccio, P.; Dragan, Y.; Ferguson, D.; Fikes, J.; et al. Early prediction of polymyxin-induced nephrotoxicity with next-generation urinary kidney injury biomarkers. Toxicol. Sci. 2014, 137, 278–291.
- Azad, M.A.K.; Roberts, K.D.; Yu, H.D.H.; Liu, B.Y.; Schofield, A.V.; James, S.A.; Howard, D.L.; Nation, R.L.; Rogers, K.; de Jonge, M.D.; et al. Significant accumulation of polymyxin in single renal tubular cells: A medicinal chemistry and triple correlative microscopy approach. Anal. Chem. 2015, 87, 1590–1595.
- Yun, B.; Azad, M.A.K.; Wang, J.; Nation, R.L.; Thompson, P.E.; Roberts, K.D.; Velkov, T.; Li, J. Imaging the distribution of polymyxins in the kidney. J. Antimicrob. Chemother. 2015, 70, 827–829.
- Velkov, T.; Yun, B.; Schneider, E.K.; Azad, M.A.; Dolezal, O.; Morris, F.C.; Nation, R.L.; Wang, J.; Chen, K.; Yu, H.H.; et al. A novel chemical biology approach for mapping of polymyxin lipopeptide antibody binding epitopes. ACS Infect. Dis. 2016, 13, 41–51.
- Manchandani, P.; Zhou, J.; Ledesma, K.R.; Truong, L.D.; Chow, D.S.; Eriksen, J.L.; Tam, V.H. Characterization of polymyxin b biodistribution and disposition in an animal model. Antimicrob. Agents Chemother. 2016, 60, 1029–1034.
- Sivanesan, S.S.; Azad, M.A.K.; Schneider, E.K.; Ahmed, M.U.; Huang, J.; Wang, J.; Li, J.; Nation, R.L.; Velkov, T. Gelofusine ameliorates colistin-induced nephrotoxicity. Antimicrob. Agents Chemother. 2017, 61, e00985-17.
- Azad, M.A.K.; Sivanesan, S.; Wang, J.; Chen, K.; Nation, R.L.; Thompson, P.E.; Roberts, K.D.; Velkov, T.; Li, J. Methionine ameliorates polymyxin-induced nephrotoxicity by attenuating cellular oxidative stress. Antimicrob. Agents Chemother. 2017, 62, e01254-17.
- Roberts, K.D.; Azad, M.A.; Wang, J.; Horne, A.S.; Thompson, P.E.; Nation, R.L.; Velkov, T.; Li, J. Antimicrobial activity and toxicity of the major lipopeptide components of polymyxin b and colistin: Last-line antibiotics against multidrug-resistant gram-negative bacteria. ACS Infect. Dis. 2015, 1, 568–575.
- Lu, X.; Chan, T.; Xu, C.; Zhu, L.; Zhou, Q.T.; Roberts, K.D.; Chan, H.K.; Li, J.; Zhou, F. Human oligopeptide transporter 2 (pept2) mediates cellular uptake of polymyxins. J. Antimicrob. Chemother. 2016, 71, 403–412.
- Dai, C.; Tang, S.; Deng, S.; Zhang, S.; Zhou, Y.; Velkov, T.; Li, J.; Xiao, X. Lycopene attenuates colistin-induced nephrotoxicity in mice via activation of the nrf2/ho-1 pathway. Antimicrob. Agents Chemother. 2015, 59, 579–585.
- Dai, C.; Li, J.; Tang, S.; Li, J.; Xiao, X. Colistin-induced nephrotoxicity in mice involves the mitochondrial, death receptor, and endoplasmic reticulum pathways. Antimicrob. Agents Chemother. 2014, 58, 4075–4085.
- Yousef, J.M.; Chen, G.; Hill, P.A.; Nation, R.L.; Li, J. Ascorbic acid protects against the nephrotoxicity and apoptosis caused by colistin and affects its pharmacokinetics. J. Antimicrob. Chemother. 2012, 67, 452–459.
- Yousef, J.M.; Chen, G.; Hill, P.A.; Nation, R.L.; Li, J. Melatonin attenuates colistin-induced nephrotoxicity in rats. Antimicrob. Agents Chemother. 2011, 55, 4044–4049.
- Ma, Z.; Wang, J.; Nation, R.L.; Li, J.; Turnidge, J.D.; Coulthard, K.; Milne, R.W. Renal disposition of colistin in the isolated perfused rat kidney. Antimicrob. Agents Chemother. 2009, 53, 2857–2864.
- Suzuki, T.; Yamaguchi, H.; Ogura, J.; Kobayashi, M.; Yamada, T.; Iseki, K. Megalin contributes to kidney accumulation and nephrotoxicity of colistin. Antimicrob. Agents Chemother. 2013, 57, 6319–6324.
- Hori, Y.; Aoki, N.; Kuwahara, S.; Hosojima, M.; Kaseda, R.; Goto, S.; Iida, T.; De, S.; Kabasawa, H.; Kaneko, R.; et al. Megalin blockade with cilastatin suppresses drug-induced nephrotoxicity. JASN 2017, 28, 1783–1791.
- Visentin, M.; Gai, Z.; Torozi, A.; Hiller, C.; Kullak-Ublick, G.A. Colistin is substrate of the carnitine/organic cation transporter 2 (octn2, slc22a5). Drug Metab. Dispos. 2017, 45, 1240–1244.
- Li, Z.D.; Luo, J.; Jia, L.H.; Wang, X.Y.; Xun, Z.K.; Liu, M. Cytochrome c suppresses renal accumulation and nephrotoxicity of polymyxin b. Hum. Exp. Toxicol. 2019, 38, 193–200.
- Azad, M.A.K.; Akter, J.; Rogers, K.L.; Nation, R.L.; Velkov, T.; Li, J. Major pathways of polymyxin-induced apoptosis in rat kidney proximal tubular cells. Antimicrob. Agents Chemother. 2015, 59, 2136–2143.
- Eadon, M.T.; Hack, B.K.; Alexander, J.J.; Xu, C.; Dolan, M.E.; Cunningham, P.N. Cell cycle arrest in a model of colistin nephrotoxicity. Physiol. Genom. 2013, 45, 877–888.
- Yun, B.; Zhang, T.; Azad, M.A.K.; Wang, J.; Nowell, C.J.; Kalitsis, P.; Velkov, T.; Hudson, D.F.; Li, J. Polymyxin b causes DNA damage in hk-2 cells and mice. Arch. Toxicol. 2018, 92, 2259–2271.
- Lee, T.W.; Bae, E.; Kim, J.H.; Jang, H.N.; Cho, H.S.; Chang, S.H.; Park, D.J. The aqueous extract of aged black garlic ameliorates colistin-induced acute kidney injury in rats. Ren. Fail. 2019, 41, 24–33.
- Jeong, B.Y.; Park, S.R.; Cho, S.; Yu, S.L.; Lee, H.Y.; Park, C.G.; Kang, J.; Jung, D.Y.; Park, M.H.; Hwang, W.M.; et al. Tgf-beta-mediated nadph oxidase 4-dependent oxidative stress promotes colistin-induced acute kidney injury. J. Antimicrob. Chemother. 2018, 73, 962–972.
- Wu, J.; Pan, X.; Fu, H.; Zheng, Y.; Dai, Y.; Yin, Y.; Chen, Q.; Hao, Q.; Bao, D.; Hou, D. Effect of curcumin on glycerol-induced acute kidney injury in rats. Sci. Rep. 2017, 7, 10114.
- He, L.; Peng, X.; Zhu, J.; Liu, G.; Chen, X.; Tang, C.; Liu, H.; Liu, F.; Peng, Y. Protective effects of curcumin on acute gentamicin-induced nephrotoxicity in rats. Can. J. Physiol. Pharm. 2015, 93, 275–282.
- Deck, L.M.; Hunsaker, L.A.; Vander Jagt, T.A.; Whalen, L.J.; Royer, R.E.; Vander Jagt, D.L. Activation of antioxidant nrf2 signaling by enone analogues of curcumin. Eur. J. Med. Chem. 2018, 143, 854–865.
- Dai, C.; Li, B.; Zhou, Y.; Li, D.; Zhang, S.; Li, H.; Xiao, X.; Tang, S. Curcumin attenuates quinocetone induced apoptosis and inflammation via the opposite modulation of nrf2/ho-1 and nf-kb pathway in human hepatocyte l02 cells. Food Chem. Toxicol. 2016, 95, 52–63.
- Klionsky, D.J.; Abdelmohsen, K.; Abe, A.; Abedin, M.J.; Abeliovich, H.; Acevedo Arozena, A.; Adachi, H.; Adams, C.M.; Adams, P.D.; Adeli, K.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12, 1–222.
- Ortega-Dominguez, B.; Aparicio-Trejo, O.E.; Garcia-Arroyo, F.E.; Leon-Contreras, J.C.; Tapia, E.; Molina-Jijon, E.; Hernandez-Pando, R.; Sanchez-Lozada, L.G.; Barrera-Oviedo, D.; Pedraza-Chaverri, J. Curcumin prevents cisplatin-induced renal alterations in mitochondrial bioenergetics and dynamic. Food Chem. Toxicol. 2017, 107, 373–385.
- Avila-Rojas, S.H.; Tapia, E.; Briones-Herrera, A.; Aparicio-Trejo, O.E.; Leon-Contreras, J.C.; Hernandez-Pando, R.; Pedraza-Chaverri, J. Curcumin prevents potassium dichromate (k2cr2o7)-induced renal hypoxia. Food Chem. Toxicol. 2018, 121, 472–482.
- Guerrero-Hue, M.; Garcia-Caballero, C.; Palomino-Antolin, A.; Rubio-Navarro, A.; Vazquez-Carballo, C.; Herencia, C.; Martin-Sanchez, D.; Farre-Alins, V.; Egea, J.; Cannata, P.; et al. Curcumin reduces renal damage associated with rhabdomyolysis by decreasing ferroptosis-mediated cell death. FASEB J. 2019, 33, 8961–8975.
- Molina-Jijon, E.; Aparicio-Trejo, O.E.; Rodriguez-Munoz, R.; Leon-Contreras, J.C.; Del Carmen Cardenas-Aguayo, M.; Medina-Campos, O.N.; Tapia, E.; Sanchez-Lozada, L.G.; Hernandez-Pando, R.; Reyes, J.L.; et al. The nephroprotection exerted by curcumin in maleate-induced renal damage is associated with decreased mitochondrial fission and autophagy. Biofactors 2016, 42, 686–702.
- Sankar, P.; Telang, A.G.; Kalaivanan, R.; Karunakaran, V.; Suresh, S.; Kesavan, M. Oral nanoparticulate curcumin combating arsenic-induced oxidative damage in kidney and brain of rats. Toxicol. Ind. Health 2016, 32, 410–421.
- Benzer, F.; Kandemir, F.M.; Kucukler, S.; Comakli, S.; Caglayan, C. Chemoprotective effects of curcumin on doxorubicin-induced nephrotoxicity in wistar rats: By modulating inflammatory cytokines, apoptosis, oxidative stress and oxidative DNA damage. Arch. Physiol. Biochem. 2018, 124, 448–457.


