 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Xiaomin Hu | + 8568 word(s) | 8568 | 2019-06-18 09:05:31 | | | |
| 2 | Bruce Ren | -4163 word(s) | 4405 | 2020-10-29 09:16:08 | | |
Video Upload Options
Kenya is among the most affected tropical countries with pathogen transmitting Culicidae vectors. For decades, insect vectors have contributed to the emergence and distribution of viral and parasitic pathogens. Outbreaks and diseases have a great impact on a country’s economy, as resources that would otherwise be used for developmental projects are redirected to curb hospitalization cases and manage outbreaks. Infected invasive mosquito species have been shown to increasingly cross both local and global boarders due to the presence of increased environmental changes, trade, and tourism. In Kenya, there have been several mosquito-borne disease outbreaks such as the recent outbreaks along the coast of Kenya, involving chikungunya and dengue. This certainly calls for the implementation of strategies aimed at strengthening integrated vector management programs. Here we look at mosquitoes of public health concern in Kenya, while highlighting the pathogens they have been linked with over the years and across various regions.
1. Introduction
The term “vector-borne” has become a commonly used term, especially in tropical and subtropical countries where emerging and re-emerging vector-related diseases frequently occur. One-sixth of human diseases is associated with vector-borne pathogens, with approximately more than half of the global population currently estimated to be in danger of contracting these diseases [1]. Hematophagous mosquitoes are the leading vectors among arthropods because of the significant role they play in disseminating microfilariae, arboviruses, and Plasmodium parasites that seem endemic to sub-Saharan Africa [2,3]. The majority of these pathogens are maintained in zoonotic cycles and humans are typically coincidental dead-end hosts with a none-to-minimal role in the cycle of the pathogen [4].
Mosquito vectoral ability is greatly influenced by the availability of conducive breeding grounds, which is in turn, influenced by the spatial heterogeneity as well as the temporal variability of the environment [5]. Mosquitoes, pathogens, and hosts each endure and reproduce within certain ideal climatic conditions and changes in these conditions can greatly alter these pathogen transmission/competences. In this scope, temperature and level of precipitation are the most influential climatic components, but other factors such as sunshine length, sea level elevation, and wind have been shown to have considerable effects [6,7]. These vectors often adjust to changes in temperature by changing topographical distribution. For instance, the advent of malaria cases in the cooler regions of East African highlands may be attributed to climate change, which has led to an increase in mosquitoes in the highlands as they warm up [8]. Variability in precipitation may also have a direct influence on distribution of mosquito-borne diseases. When precipitation increases, the presence of disease vectors is also expected to rise due to the expansion of the existent larval habitat and emergence of new breeding zones [6]. Each mosquito species has unique environmental resilience limits dependent upon the availability of favorable aquatic larval habitats and the closeness of vertebrate hosts that serve as their source of blood meals. This reliance of mosquito species on aquatic environments is a constant part of their lifecycle and the availability of a suitable aquatic domain, which is a requirement for the development of eggs, larvae, and pupae, and basically determines the abundance of mosquito species.
Kenya represents a topographically diverse tropical/subtropical country which harbors a large diversity of mosquito species of public health importance. Many factors contribute to the extensive proliferation of mosquitoes ranging from global warming, sporadic floods, improper waste disposal, irrigation canals, presence of several lakes/rivers, and low altitudes around coastal regions. Consequently, an upsurge in emerging and re-emerging mosquito-borne pathogens over the years has been recorded with increased research efforts geared towards mosquitoes and their pathogens [9]. Increased urbanization, tourism, and international trade have led some of these species and pathogens to cross local and international borders to new territories. This dispersal poses both local and global health threats if proper mitigation measures are not put in place. Nevertheless, not all mosquito species are associated with human diseases, thus, this review highlights species of the main mosquito genera to which pathogens have been associated/detected and their countrywide distribution based on published data and reported cases. Additionally, this information provides a guide to proper mosquito control strategies by comparing the methods currently applied in the country and proposed alternative methods applied in other countries affected by mosquito disease burden.
2. Mosquito-Borne Disease Endemic Regions in Kenya
Kenya, as a tropical country, is among the most affected sub-Saharan regions with mosquito-related ailments. The countrywide distribution of mosquito species is, however, not well documented with most studies focusing on disease endemic regions. Some of these regions include Garissa, Mandera, and Turkana situated in the north and north-eastern parts of the country and characterized as arid and semi-arid regions (Figure 1A). During rainy seasons, flood water acts as breeding areas for mosquito species. Rift Valley fever outbreaks were recorded in this area in 1997/98 and 2006/07 [10,11]. Other viruses reported from the regions include West Nile virus (WNV), Ndumu virus (NDUV), Babanki virus (BBKV), and orthobunyaviruses isolated from the flood water Aedes and Culex species [4]. For decades, the coastal region of Kenya (Kwale, Kilifi, Mombasa, Lamu) has also reported multiple outbreaks resulting from mosquito-borne pathogens making it one of the most endemic regions. The contributing factors are majorly the low altitude which provides a conductive environment for mosquito breeding and high human population composed of both locals and global tourists. Mosquitoes of interest in this coastal region in terms of pathogen transmission include: Aedes aegypti that transmits dengue fever and chikungunya [12,13]; Anopheles species (Anopheles gambiae, Anopheles arabiensis, Anopheles funestus, Anopheles merus) which are associated with malaria and bancroftian filariasis sporozoites [14,15], and Culex quinquefasciatus [16] among others as demonstrated in Figure 1A–C.
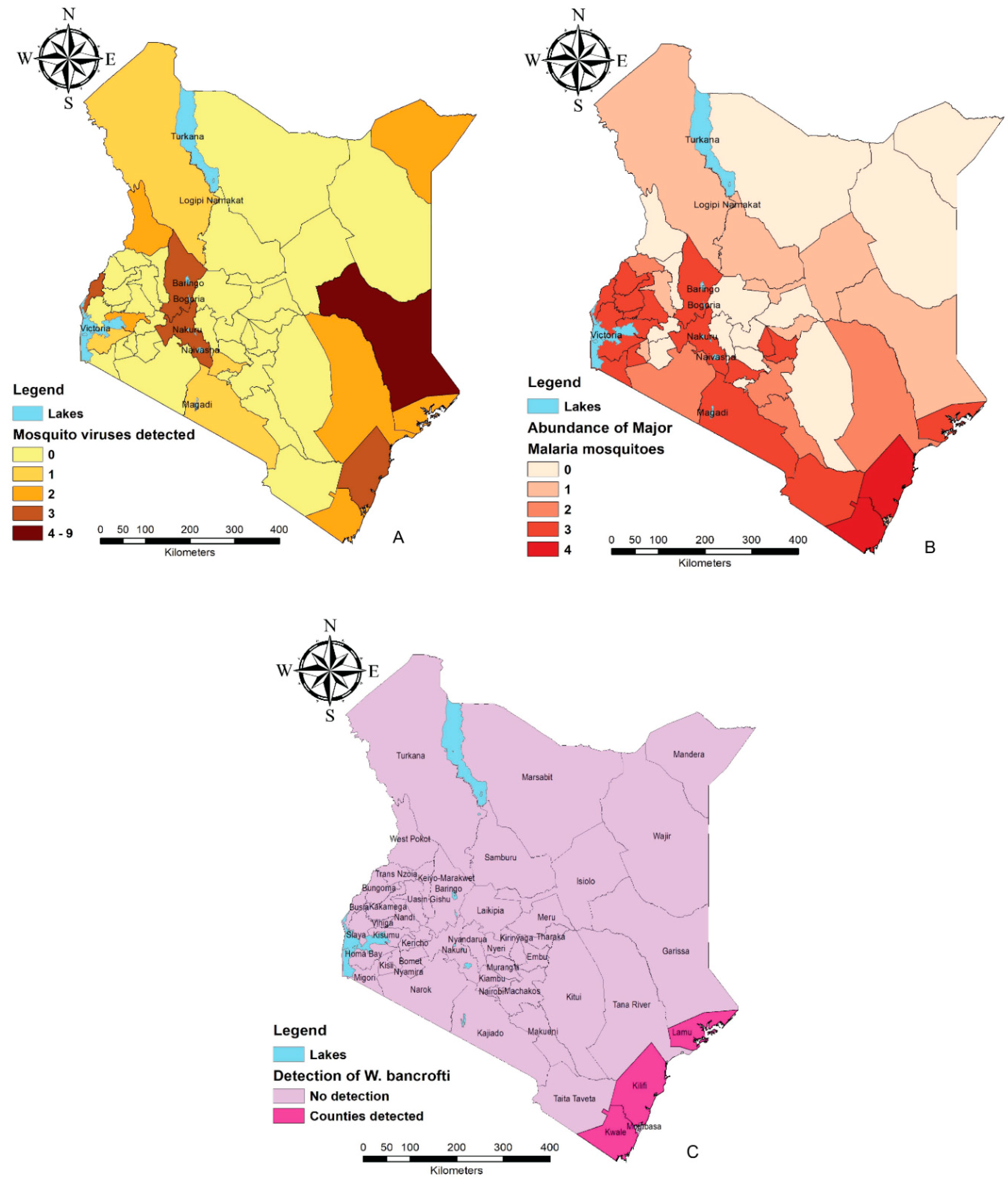
Figure 1. Mosquito-borne disease endemic regions based on distribution of pathogens and associated mosquito species (maps were constructed using the free and open-source Quantum GIS software (https://qgis.org/en/site/) using data compiled from Table 1 and Table 2. (A) Abundance of mosquito-borne viruses detected/isolated in various counties. (B) Distribution of the major malaria vectors in numbers in different counties. (C) Counties in which Wuchereria bancrofti has been detected from mosquitoes.
Table 1. Summary of the main mosquito species in which viruses have been detected/isolated in Kenya.
|
Genera |
Species |
Virus Isolated/Detected 1 |
County of Virus Detection |
Reference |
|---|---|---|---|---|
|
Aedes |
A. aegypti |
DENV, CHKV |
Mombasa, Mandera, Kilifi, Lamu, Busia |
|
|
A. africanus |
YFV |
Baringo, |
[59] |
|
|
A. albicosta |
DENV, CHKV |
Mombasa, Kilifi, Lamu, Kwale |
[33] |
|
|
A. circumluteolus |
RVFV, BBKV, NDUV, SMFV |
Garissa |
||
|
A. fryeri |
DENV |
Mombasa, Kilifi, Lamu, Kwale |
[33] |
|
|
A. fulgens |
DENV, CHKV |
Mombasa, Kilifi, Lamu, Kwale |
[33] |
|
|
A. keniensis |
YFV |
Baringo |
[59] |
|
|
A. Luridus |
NDUV |
Tana River |
[4] |
|
|
A. mcintoshi |
RVFV, NDUV, PGAV, BUNV, BBKV, PGAV, SMFV, NRIV |
Garissa |
||
|
|
DENV, CHKV |
Mombasa, Kilifi, Lamu, Kwale |
[33] |
|
|
A. ochraceus |
RVFV, NDUV, BUNV, BBKV, SNBV, SMFV |
Garissa |
||
|
|
DENV, CHKV |
Mombasa, Kilifi, Lamu, Kwale |
[33] |
|
|
A. pembaensis |
RVFV |
Kilifi |
[4] |
|
|
|
DENV, CHKV |
Mombasa, Kilifi, Lamu, Kwale |
[33] |
|
|
A. sudanensis |
BBKV, SNBV, WNV |
Garissa |
[36] |
|
|
|
|
NDUV |
Tana River |
[39] |
|
Anopheles |
An. funestus |
ONNV |
Kisumu |
[59] |
|
|
BUNV |
Kajiado |
[4] |
|
|
|
NRIV |
Tana River |
[4] |
|
|
An. gambiae |
BUNV |
Homabay |
[20] |
|
|
An. squamosus |
RVFV |
Garissa |
[11] |
|
|
Culex |
Cx. bitaeniorhynchus |
RVFV |
Kilifi |
[11] |
|
|
NDUV |
Tana River |
[39] |
|
|
Cx. cinereus |
NDUV |
Busia |
[4] |
|
|
Cx. pipiens |
USUV |
Kisumu |
[4] |
|
|
|
NDUV |
Garissa, Tana River |
||
|
Cx. poicilipes |
RVFV |
Kilifi |
[11] |
|
|
Cx. quinquefasciatus |
RVFV |
Baringo, Garissa |
||
|
|
WNV, SNBV |
Garissa |
||
|
Cx. rubinotus |
NDUV |
Baringo |
[4] |
|
|
Cx. univittatus |
RVFV |
Baringo |
[11] |
|
|
|
BUNV |
Homa Bay |
[20] |
|
|
|
SNBV |
West Pokot, Nakuru, Busia |
||
|
|
WNV |
Garissa, Turkana, West Pokot |
||
|
Cx. vansomereni |
NDUV |
Tana River |
[39] |
|
|
|
BBKV, SNBV |
Nakuru |
||
|
Cx. zombaensis |
RVFV |
Nakuru |
[62] |
|
|
|
BBKV |
Kiambu |
[4] |
|
|
Mansonia |
Mn. africana |
RVFV |
Nakuru, Baringo, Garissa |
|
|
|
NDUV |
Baringo |
[4] |
|
|
Mn. uniformis |
RVFV |
Baringo, Garissa |
||
|
|
NDUV |
Baringo |
[36] |
Genera: A, Aedes; An, Anopheles; Cx, Culex; Mn, Mansonia. 1 Abbreviations: DENV—dengue virus, CHKV—chikungunya virus, RVFV—Rift Valley fever virus, NDUV—Ndumu virus, PGAV—Pongola virus, BUNV—Bunyamwera virus, BBKV—Babanki virus, SMFV—Semliki Forest virus, NRIV—Ngari virus, YFV—yellow fever virus, SNBV—Sindbis virus, WNV—West Nile virus, USUV—Usutu virus, ONNV—O’nyong’nyong virus.
Table 2. Summary of the dominant malaria and Bancroftian filariasis mosquito vectors distributed in Kenya.
|
Parasite |
Associated Human Ailment |
Dominant Mosquito Species |
Counties of Vector Distribution |
Reference |
|---|---|---|---|---|
|
Plasmodium falciparum |
Malaria |
An. gambiae s.s., An. arabiensis, An. funestus, An. merus |
Kwale, Kilifi |
|
|
An. gambiae ss, An. arabiensis, An. funestus |
Taita-Taveta, Lamu, Kajiado, Embu, Nakuru, Baringo, Bungoma, Kirinyaga, Kiambu, Busia, Siaya, Kakamega, Vihiga, Homabay, Migori, Kisii, Kisumu, Nandi |
|||
|
An. gambiae ss, An. arabiensis |
Narok |
|||
|
An. arabiensis, An. funestus |
Tana-River, Makueni, Machakos, Trans-Nzoia |
|||
|
An. funestus |
Samburu, Isiolo, Garissa, Mombasa, Uasin-Gishu, Nyamira |
|||
|
An. arabiensis |
Turkana |
|||
|
An. gambiae ss |
Tharaka-Nithi |
|||
|
Wuchereria bancrofti |
Bancroftian filariasis |
An. gambiae sl, An. funestus, Cx. quinquefasciatus |
Kwale, Kilifi, Lamu |
Genera: An, Anopheles; Cx, Culex.
3. Overview of the Main Mosquito Genera of Public Health Importance in Kenya and Their Geographical Distribution
The main mosquito species associated with human etiologies in the country belong to four genera (Aedes, Anopheles, Culex, and Mansonia) as discussed below. Their distribution in Kenya has mainly been studied in disease endemic regions and as a result of outbreaks. These data are summarized in Table 1 and Table 2.
3.1. Aedes
3.2. Anopheles
3.3. Culex
3.4. Mansonia
The Mansonia mosquito species belong to the sub-family Culicinae and are normally characterized by their large body size and asymmetrical wing scale structure with sparkling on their wing veins and legs. They show some resemblance with Aedes, Culex, and Coquillettidia mosquito but the simplicity of their tarsal claws in structure and a truncated abdomen in females sets it apart [113]. Furthermore, Mansonia preferably breed in ponds and permanent waters with aquatic plants where larvae can burrow into the dead plants at the bottom or cling to the roots of the live ones [114]. Its genera comprise 25 globally distributed species in two subgenera: Mansonioides (10 species) and Mansonia (15 species).
The distribution of these species comes with increased vectorial role of human etiologies. Mansonia titillans, a common species in the Americas, is a known vector of Venezuelan equine encephalitis [115]. Recently, Argentina detected SLEV and BUNV for the first time from this species, proving its competent nature to transmit the viruses [116]. In south-east Asia, Mn. bonneae and Mn. dives have been linked to various cases of Brugian and lymphatic filariasis [117]. Africa, as one of the most affected regions of vector-borne diseases, harbors two main and probably the most common Mansonia spp. (Mn. africana and Mn. uniformis). These species have been reported to vector W. bancrofti’s lymphatic filariasis [118,119], RVF [34], Zika [120], among other diseases. Kenya is not an exception, as studies have shown that the two Mansonia spp. are distributed within the country and harbor pathogens of human health importance [4,11,36,40,60].
LaBeaud et al. [60] showed that Mansonia mosquitoes could have significantly contributed to the vectorial transmission of RVF in the 2006–2007 outbreak in the north-eastern region. Out of the 12,080 mosquitos collected during the study, 682 were identified to be Mansonia spp. Further investigation based on molecular screening identified three out of eight pools of Mansonia spp. to be positive for RVFV. Mansonia, like many other female mosquitoes, feed on vertebrate blood for egg development. These vertebrates can lead to amplification of a disease in case of outbreaks. Virus screening from blood-fed Mansonia spp. by Lutomiah et al. [40] during this same RVF outbreak, showed that goats and sheep were the greatest amplifiers of the virus.
Owing to the fact that Mn. uniformis and Mn. africana preferably breed around permanent water bodies, flooded lagoons, and swamps, they have been shown to dominate lake regions in the country. A study by Lutomiah et al. [22] on the abundance of mosquito vectors in various ecological zones in the country revealed high abundance of Mansonia spp. in Baringo (71%) and Kisumu (23%), the home places of Lake Baringo and Lake Victoria. Interestingly, it is here (Baringo) that positive strains of RVFV and NDUV had earlier been detected in Mn. uniformis and Mn. africana [11,36]. This was further evidenced by Ajamma et al. [18], where these species dominated in abundance in the islands and mainlands of both Lake Victoria and Lake Baringo.
Even though Zika virus and W. bancrofti have been detected from Mansonia spp. in some African states [118,120], this has not been the case in Kenya. In a study at the Kenyan Tana delta on vectorial potential of Mansonia spp. to transmit W. bancrofti; all 236 species collected tested negative on PCR (polymerase chain reaction) analysis [121]. However, the study recommended further assessment to be done by infecting Mansonia spp. experimentally with W. bancrofti microfilaria to identify whether or not they can support development of microfilaria up to the infective stages.
References
- World Health Organization. Global Vector Control Response 2017–2030. 2017. Available online: https://apps.who.int/iris/bitstream/handle/10665/259205/9789241512978-eng.pdf;jsessionid=9C11656832A16B1404639714E44BF08B?sequence=1 (accessed on 4 March 2019)
- Braack, L.; Gouveia De Almeida, A.P.; Cornel, A.J.; Swanepoel, R.; De Jager, C. Mosquito-borne arboviruses of African origin: Review of key viruses and vectors. Parasites Vectors 2018, 11, 29, doi:1186/s13071-017-2559-9.
- Centers for Disease Control. Lymphatic Filariasis: Epidemiology and Risk Factors, 2019. Available online: https://www.cdc.gov/parasites/lymphaticfilariasis/epi.html (accessed on 4 March 2019).
- Ochieng, C.; Lutomiah, J.; Makio, A.; Koka, H.; Chepkorir, E.; Yalwala, S.; Mutisya, J.; Musila, L.; Khamadi, S.; Richardson, J.; et al. Mosquito-borne arbovirus surveillance at selected sites in diverse ecological zones of Kenya; 2007–2012. J. 2013, 10, 140, doi:10.1186/1743-422X-10-140.
- Altizer, S.; Dobson, A.; Hosseini, P.; Hudson, P.; Pascual, M.; Rohani, P. Seasonality and the dynamics of infectious diseases. Lett. 2006, 9, 467–484, doi:10.1111/j.1461-0248.2005.00879.x.
- Patz,A.; Githeko, A.K.; McCarty, J.P.; Hussein, S.; Confalonieri, U.; DeWet, N. Climate change and Infectious Diseases. World Health Organization. 2003. Available online: https://www.who.int/globalchange/publications/
- climatechangechap6.pdf (accessed on 4 March 2019).
- Shi, Z. Impact of Climate Change on the Global Environment and Associated Human Health. Res. 2018, 5, e4934, doi:10.4236/oalib.1104934.
- Himeidan, Y.E.; Kweka, E.J. Malaria in East African highlands during the past 30 years: Impact of environmental changes. Physiol. 2012, 3, 315, doi:10.3389/fphys.2012.00315.
- Owino, E.A. Aedes mosquitoes and emerging neglected diseases of Kenya. Int. J. Mosq. Res. 2018, 5, 5, 2348–5906
- Woods, C.W.; Karpati, A.M.; Grein, T. McCarthy, N.; Gaturuku, P.; Muchiri, E.; Dunster, L.; Henderson, A.; Khan, A.S.; Al, R.S. An Outbreak of Rift Valley Fever in Northeastern Kenya, 1997–98. Infect. Dis. 2002, 8, 138–144, doi:10.3201/eid0802.010023.
- Sang, R.; Kioko, E.; Lutomiah, J.; Warigia, M.; Ochieng, C.; O’Guinn, M.; Lee, J.S.; Koka, H.; Godsey, M.; Hoel, D.; et al. Rift Valley fever virus epidemic in Kenya, 2006/2007: The entomologic investigations. J. Trop. Med. Hyg. 2010, 83, 28–37, doi:10.4269/ajtmh.2010.09-0319.
- Lutomiah, J.; Barrera, R.; Makio, A.; Mutisya, J.; Koka, H.; Owaka, S.; Koskei, E.; Nyunja, A.; Eyase, F.; Coldren, R.; et al. Dengue Outbreak in Mombasa City, Kenya, 2013–2014: Entomologic Investigations. PLoS Negl. Trop. Dis. 2016, 10, e0004981, doi:10.1371/journal.pntd.0004981.
- World Health Organization. Chikungunya—Mombasa, Kenya. 2018. Available online: https://www.who.int/
- csr/don/27-february-2018-chikungunya-kenya/en/ (accessed on 10 March 2019).
- Mwangangi, J.M.; Midega, J.; Kahindi, S.; Njoroge, L.; Nzovu, J.; Githure, J.; Mbogo, C.M.; Beier, J.C. Mosquito species abundance and diversity in Malindi, Kenya and their potential implication in pathogen transmission. Res. 2012, 110, 61–71, doi:10.1007/s00436-011-2449-6.
- Ogola, E.O.; Odero, J.O.; Mwangangi, J.M.; Masiga, D.K.; Tchouassi, D.P. Population genetics of Anopheles funestus, the African malaria vector, Kenya. Parasites Vectors 2019, 12, 15, doi:10.1186/s13071-018-3252-3.
- Atoni, E.; Wang, Y.; Karungu, S.; Waruhiu, C.; Zohaib, A.; Obanda, V.; Agwanda, B.; Mutua, M.; Xia, H.; Yuan, Z. Metagenomic virome analysis of Culex mosquitoes from Kenya and China. Viruses 2018, 10, 30, doi:10.3390/v10010030.
- Minakawa, N.; Dida, G.O.; Sonye, G.O.; Futami, K.; Njenga, S.M. Malaria vectors in Lake Victoria and adjacent habitats in western Kenya. PLoS ONE 2012, 7, e32725, doi:10.1371/journal.pone.0032725.
- Ajamma, Y.K.; Villinger, J.; Omondi, D.; Salifu, D.; Onchuru,T.O.; Njoroge, L.; Muigai, A.W.T.; Masiga, D.K. Composition and genetic diversity of mosquitoes (Diptera: Culicidae) on Islands and mainland shores of Kenya’s lakes Victoria and baringo. Med. Entomol. 2016, 53, 1348–1363, doi:10.1093/jme/tjw102.
- Ondiba, I.M.; Oyieke, F.A.; Nyamongo, I.K.; Estambale, B.B. Diversity, distribution and abundance of potential rift valley fever vectors in Baringo County, Kenya. J. Mosq. Res. 2017, 4, 42–48.
- Ajamma, Y.U.; Onchuru, T.O.; Ouso, D.O.; Omondi, D.; Masiga, D.K.; Villinger, J. Vertical transmission of naturally occurring Bunyamwera and insect-specific flavivirus infections in mosquitoes from islands and mainland shores of Lakes Victoria and Baringo in Kenya. PLoS Negl. Trop. Dis. 2018, 12, e0006949, doi:10.1371/journal.pntd.0006949.
- Ofulla, A.V.O.; Karanja, D.; Omondi, R.; Okurut, T.; Matano, A.; Jembe, T.; Abila, R.; Boera, P.; Gichuki, J. Relative abundance of mosquitoes and snails associated with water hyacinth and hippo grass in the Nyanza gulf of Lake Victoria. Lakes Reserv. Res. Manag. 2010, 15, 225–271, doi:1111/j.1440-1770.2010.00434.x.
- Lutomiah, J.; Bast, J.; Clark, J.; Richardson, J.; Yalwala, S.; Oullo, D.; Mutisya, J.; Mulwa, F.; Musila, L.; Khamadi, S.; et al. Abundance, diversity, and distribution of mosquito vectors in selected ecological regions of Kenya: Public health implications. Vector Ecol. 2013, 38, 134–142, doi:10.1111/j.1948-7134.2013.1201.
- Turell, M.J.; Lee, J.S.; Richardson, J.H.; Sang, R.C.; Kioko, E.N.; Agawo, M.O.; Pecor, J.; O’guinn, M.L. Vector Competence of Kenyan Culex zombaensis And Culex quinquefasciatus mosquitoes for Rift Valley Fever Virus. Am. Mosq. Control Assoc. 2007, 23, 378–382, doi:10.2987/5645.1.
- Britch, S.C.; Binepal, Y.S.; Ruder, M.G.; Kariithi, H.M.; Linthicum, K.J.; Anyamba, A.; Small, J.L.; Tucker, C.J.; Ateya, L.O.; Oriko, A.A.; et al. Rift Valley Fever Risk Map Model and Seroprevalence in Selected Wild Ungulates and Camels from Kenya. PLoS ONE 2013, 8, e66626, doi:10.1371/journal.pone.0066626.
- Tuno, N.; Okeka, W.; Minakawa, N.; Takagi, M.; Yan, G. Survivorship of Anopheles gambiae sensu stricto (Diptera: Culicidae) Larvae in Western Kenya Highland Forest. Med. Entomol. 2005, 42, 270–277.doi:10.1093/jmedent/42.3.270.
- Doucoure, S.; Drame, P.M. Salivary biomarkers in the control of mosquito-borne diseases. Insects 2015, 6, 961–976, doi:10.3390/insects6040961.
- Iwashita, H.; Higa, Y.; Futami, K.; Lutiali, P.A.; Njenga, S.M.; Nabeshima, T.; Minakawa, N. Mosquito arbovirus survey in selected areas of Kenya: Detection of insect-specific virus. Med. Health 2018, 46, doi:10.1186/s41182-018-0095-8.
- Bravo, L.; Roque, V.G.; Brett, J.; Dizon, R.; L’Azou, M. Epidemiology of dengue disease in the Philippines (2000–2011): A systematic literature review. PLoS Negl. Trop. Dis. 2014, 8, e3027, doi:10.1371/journal.pntd.0003027.
- Musso, D.; Rodriguez-Morales, A.J.; Levi, J.E.; Cao-Lormeau, V.M.; Gubler, D.J. Unexpected outbreaks of arbovirus infections: Lessons learned from the Pacific and tropical America. Lancet Infect. Dis. 2018, 18, e355–e361, doi:10.1016/ S1473-3099(18)30269-X.
- Carrington, L.B.; Simmons, C.P. Human to mosquito transmission of dengue viruses. Immunol. 2014, 5, 290, doi:10.3389/fimmu.2014.00290.
- Sharp, T.M.; Mackay, A.J.; Santiago, G.A.; Hunsperger, E.; Nilles, E.J.; Perez-Padilla, J.; Tikomaidraubuta, K.S.; Colon, C.; Amador, M.; Chen, T.; et al. Characteristics of a dengue outbreak in a remote Pacific Island chain—Republic of the Marshall Islands, 2011–2012. PLoS ONE 2014, 9, e108445, doi:10.1371/journal.pone.0108445.
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.N.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Bortel, W.V. et al. The global distribution of the arbovirus vectors Aedes aegypti and albopictus. elife 2015, 4, e08347, doi:10.7554/eLife.08347.
- Ngala, J.C.; Schmidt Chanasit, J. Entomological Coinfections of Arboviruses Dengue and Chikungunya Viruses along the Coastline of Kenya. Mosq. Res. 2018, 8, 1, doi:10.5376/jmr.2018.08.0001.
- Arum, S.O.; Weldon, C.W.; Orindi, B.; Landmann, T.; Tchouassi, D.P.; Affognon, H.D.; Sang, R. Distribution and diversity of the vectors of Rift Valley fever along the livestock movement routes in the northeastern and coastal regions of Kenya. Parasites Vectors 2015, 8, 294, doi:10.1186/s13071-015-0907-1.
- Tchouassi, D.P.; Bastos, A.D.S.; Sole, C.L.; Diallo, M.; Lutomiah, J.; Mutisya, J.; Mulwa, F.; Borgemeister, C.; Sang, R.; Torto, B.; et al. Population Genetics of Two Key Mosquito Vectors of Rift Valley Fever Virus Reveals New Insights into the Changing Disease Outbreak Patterns in Kenya. PLoS Negl. Trop. Dis. 2014, 8, e3364 , doi:10.1371/journal.pntd.0003364.
- Crabtree, M.; Sang, R.; Lutomiah, L.; Richardson, J.; Miller, B. Arbovirus surveillance of mosquitoes collected at sites of active Rift Valley fever virus transmission: Kenya, 2006– J. Med. Entomol. 2009, 46, 961–964, doi:10.1603/033.046.0431.
- Sigei, F.; Nindo, F.; Mukunzi, S.; Ng’ang’a, Z.; Sang, R. Evolutionary analyses of Sindbis virus strains isolated from mosquitoes in Kenya. Virol. 2018, 163, 2465–2469, doi:10.1007/s00705-018-3869-8.
- Lutomiah, J.; Ongus, J.; Linthicum, K.J.; Sang, R. Natural Vertical Transmission of Ndumu Virus in Culex pipiens (Diptera: Culicidae) Mosquitoes Collected as Larvae. Med. Entomol. 2014, 51, 1091–1095, doi:10.1603/me14064.
- Sang, R.; Lutomiah, J.; Said, M.; Makio, A.; Koka, H.; Koskei, E.; Nyunja, A.; Owaka, S.; Matoke-Muhia, D.; Bukachi, S.; et al. Effects of irrigation and rainfall on the population dynamics of rift valley fever and other arbovirus mosquito vectors in the epidemic-prone Tana-River County, Kenya. Med. Entomol. 2017, 54, 460–470, doi:10.1093/jme/tjw206.
- Lutomiah, J.; Omondi, D.; Masiga, M.; Mutai, C.; Mireji, P.O.; Ongus, J.; Linthicum, K.J.; Sang, R. Blood Meal Analysis and Virus Detection in Blood-Fed Mosquitoes Collected During the 2006–2007 Rift Valley Fever Outbreak in Kenya. Vector-Borne Zoonotic Dis. 2014, 14, 656–664, doi:10.1089/vbz.2013.1564.
- Sanders, E.J.; Marfin, A.A.; Tukei, P.M.; Kuria, G.; Ademba, G.; Agata, N.N.; Ouma, J.O.; Cropp, C.B.; Karabatsos, N.; Reiter, P.; et al. First recorded outbreak of yellow fever in Kenya, 1992–1993. II. Entomologic investigations. J. Trop. Med. Hyg. 1998, 59, 644–649.
- Ellis, B.R.; Sang, R.C.; Horne, K.M.; Higgs, S.; Wesson, D.M. Yellow fever virus susceptibility of two mosquito vectors from Kenya, East Africa. R. Soc. Trop. Med. Hyg. 2012, 106, 387–389, doi:10.1016/j.trstmh.2012.02.007.
- Mulwa, F.; Lutomiah, J.; Chepkorir, E.; Okello, S.; Eyase, F.; Tigoi, C.; Kahato, M.; Sang, R. Vector competence of Aedes bromeliae and Aedes vitattus mosquito populations from Kenya for chikungunya virus. PLoS Negl. Trop. Dis. 2018, 12, doi:10.1371/journal.pntd.0006746.
- Sang, R.; Arum, S.; Chepkorir, E.; Mosomtai, G.; Tigoi, C.; Sigei, F.; Lwande, O.W.; Landmann, T.; Affognon, H.; Ahlm, C.; et al. Distribution and abundance of key vectors of Rift Valley fever and other arboviruses in two ecologically distinct counties in Kenya. PLoS Negl. Trop. Dis. 2017, 11, e0005341, doi:10.1371/journal.pntd.0005341.
- Powell, J.; Tabachnick, W. History of domestication and spread of Aedes aegypti—A Review. Memórias do Instituto Oswaldo Cruz 2013, 108, 11–17, doi:10.1590/0074-0276130395.
- Yalwala, S.; Clark, J.; Oullo, D.; Ngonga, D.; Abuom, D.; Wanja, E.; Bast, J. Comparative efficacy of existing surveillance tools for Aedes aegypti in Western Kenya. Vector Ecol. 2015, 40, 301–307, doi:10.1111/jvec.12168.
- Ndenga, B.A.;, Mutuku, F.M.; Ngugi, H.N.; Mbakaya, J.O.; Aswani, P.; Musunzaji, P.S.; Vulule, J.; Mukoko, D.; Kitron, U.; LaBeaud, A.D. Characteristics of Aedes aegypti adult mosquitoes in rural and urban areas of western and coastal Kenya. PLoS ONE 2017, 12, e0189971, doi:10.1371/journal.pone.0189971.
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B.; Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222–227, doi:10.1038/nature13964.
- Monath, T. Dengue: The risk to developed and developing countries. Natl. Acad. Sci. USA 1994, 91, 2395–2400.
- Ngugi, H.N.; Mutuku, F.M.; Ndenga, B.A.; Musunzaji, P.S.; Mbakaya, J.O.; Aswani, P.; Irungu, L.W.; Mukoko, D.; Vulule, J.; Kitron, U.; et al. Characterization and productivity profiles of Aedes aegypti (L.) breeding habitats across rural and urban landscapes in western and coastal Kenya. Parasites Vectors 2017, 10, 331, doi:10.1186/s13071-017-2271-9.
- Johnson, B.; Musoke, S.; Ocheng, D.; Gichogo, A.; Rees, P. Dengue-2 Virus In Kenya. Lancet 1982, 320, 208–209.
- Obonyo, M.; Fidhow, A.; Ofula, V. Investigation of laboratory confirmed dengue outbreak in north-eastern Kenya, 2011. PLoS ONE 2018, 13, e0198556, doi:10.1371/journal. pone.0198556.
- Voice of Africa. Kenya Health Officials Issue Alert Over Dengue Fever Outbreak. 2017. Available online: https://www.voanews.com/a/kenya-health-officials-alert-dengue-fever-outbreak/3843040.html (accessed on 25 March 2019).
- World Health Organization. Rift Valley fever—Kenya. 2018. Available online: https://www.who.int/csr/don/
- 18-june-2018-rift-valley-fever-kenya/en/ (accessed on 25 March 2019).
- Sergon, K.; Njuguna, C.; Kalani, R.; Ofula, V.; Onyango, C.; Konongoi, L.S.; Bedno, S.; Burke, H.; Dumilla, A.M.; Konde, J.; et al. Seroprevalence of Chikungunya Virus (CHIKV) Infection on Lamu Island, Kenya, October 2004. J. Trop. Med. Hyg. 2008, 78, 333–337, doi:10.4269/ajtmh.2008.78.333.
- Berry, I.M.; Eyase, F.; Pollett, S.; Konongoi, S.L.; Joyce, M.G.; Figueroa, K.; Ofula, V.; Koka, H.; Koskei, E.; Nyunja, A.; et al. Global Outbreaks and Origins of a Chikungunya Virus Variant Carrying Mutations Which May Increase Fitness for Aedes aegypti: Revelations from the 2016 Mandera, Kenya Outbreak. J. Trop. Med. Hyg. 2019, 100, 1249-1257, doi:10.4269/ajtmh.18-0980.
- Konongoi, S.L.; Ofula, V.; Nyunja, A.; Owaka, S.; Koka, H.; Makio, A.; Koskei, E.; Eyase, F.; Langat, D.; Schoepp, R.J.; et al. Detection of dengue virus serotypes 1, 2 and 3 in selected regions of Kenya: 2011–2014. J. 2016, 13, 182, doi:10.1186/s12985-016-0641-0.
- Konongoi, S.L.; Nyunja, A.; Ofula, V.; Owaka, S.; Koka, H.; Koskei, E.; Eyase, F.; Langat, D.; Mancuso, J.; Lutomiah, J.; et al. Human and entomologic investigations of chikungunya outbreak in Mandera, Northeastern Kenya, 2016. PLoS ONE 2018, 13, e0205058, doi:10.1371/journal.pone.0205058.
- Johnson, B.K.; Gichogo, A.; Gitau, G.; Patel, N.; Ademba, G.; Kirui, R.; Highton, R.B.; Smith, D.H. Recovery of o’nyong-nyong virus from Anopheles funestus in Western Kenya. R. Soc. Trop. Med. Hyg. 1981, 75, 239–241.
- LaBeaud, A.D.; Sutherland, L.J.; Muiruri, S.; Muchiri, E.M.; Gray, L.R.; Zimmerman, P.A.; Hise, A.G.; King, C.H. Arbovirus prevalence in mosquitoes, Kenya. Infect. Dis. 2011, 17, 233–241, doi:10.3201/eid1702.091666.
- Miller, B.R.; Nasci, R.S.; Godsey, M.S.; Savage, H.M.; Lutwama, J.J.; Lanciotti, R.S.; Peters, C.J. First field evidence for natural vertical transmission of West Nile virus in Culex univittatus complex mosquitoes from Rift Valley province, Kenya. J. Trop. Med. Hyg. 2000, 62, 240–246, doi:10.4269/ajtmh.2000.62.240.
- Logan, T.M.; Linthicum, K.J.; Ksiazek, T.G. Isolation of Rift Valley fever virus from mosquitoes collected during an outbreak in domestic animals in Kenya. Med. Entomol. 1992, 28, 293–295.
- Centers for Disease Control. Anopheles 2015. Available online: https://www.cdc.gov/malaria/
- about/biology/index.html#tabs-1-5 (accessed on 25 March 2019).
- Harbach, R.E. The Culicidae (Diptera): A review of taxonomy, classification and phylogeny. Zootaxa 2007, 1668, 591–638, doi:10.11646/zootaxa.1668.1.28.
- World Health Organization. World Malaria Report at Glance. 2018. Available online: https://www.who.int/
- malaria/media/world-malaria-report-2018/en/ (accessed on 25 March 2019).
- Afrane, Y.A.; Bonizzoni, M.; Yan, G. Secondary Malaria Vectors of Sub-Saharan Africa: Threat to Malaria Elimination on the Continent. IntechOpen 2016, 20, 473–490, doi:10.5772/65359.
- Mouchet, J.; Carnevale, P.; Coosemans, M.; Julvez, J.; Manguin, S.; Richard-Lenoble, D.; Sircoulon, J. Biodiversity of Malaria Worldwide; John Libbey Eurotext: Montrouge, France, 2001.
- Coetzee, M.; Hunt, R.H.; Wilkerson, R.; Della Torre, A.; Coulibaly, M.B.; Besansky, N.J. Anopheles coluzzii and Anopheles amharicus, new members of the Anopheles gambiae Zootaxa 2013, 3619, 246–274, doi:10.11646/zootaxa.3619.3.2.
- Kibret, S.; Wilson, G.G.; Tekie, H.; Petros, B. Increased malaria transmission around irrigation schemes in Ethiopia and the potential of canal water management for malaria vector control. J. 2014, 13, 360, doi:10.1186/1475-2875-13-360.
- Lobo, N.F.; St Laurent, B.; Sikaala, C.H.; Hamainza, B.; Chanda, J.; Chinula, D.; Krishnankutty, S.M.; Mueller, J.D.; Deason, N.A.; Hoang, Q.T.; et al. Unexpected diversity of Anopheles species in Eastern Zambia: Implications for evaluating vector behavior and interventions using molecular tools. Rep. 2015, 5, 17952,doi:10.1038/srep17952.
- Okiro, E.A.; Alegana, V.A.; Noor, A.M.; Snow, R.W. Changing malaria intervention coverage, transmission and hospitalization in Kenya. J. 2010, 9, 285, doi:10.1186/1475-2875-9-285.
- Centers for Disease Control. CDC Activities in Kenya. 2018. Available online: https://www.cdc.gov/malaria/
- malaria_worldwide/cdc_activities/kenya.html (accessed on 25 March 2019).
- Macharia, P.M.; Giorgi, E.; Noor, A.M.; Waqo, E.; Kiptui4, R.; Okiro, E.A.; Snow, R.W. Spatio-temporal analysis of Plasmodium falciparum prevalence to understand the past and chart the future of malaria control in Kenya. J. 2018, 17, 340, doi:10.1186/s12936-018-2489-9.
- Okara, R.M.; Sinka, M.E.; Minakawa, N.; Mbogo, C.M.; Hay, S.I.; Snow, R.W. Distribution of the main malaria vectors in Kenya. J. 2010, 9, 69, doi:10.1186/1475-2875-9-69.
- Olanga, E.A.; Okombo, L.; Irungu, L.W.; Mukabana, W.R. Parasites and vectors of malaria on Rusinga Island, Western Kenya. Parasites Vectors 2015, 8, 250, doi:10.1186/s13071-015-0860-z.
- Ototo, E.N.; Mbugi, J.P.; Wanjala, C.L.; Zhou, G.; Githeko, A.K.; Yan, G. Surveillance of malaria vector population density and biting behaviour in western Kenya. J. 2015, 14, 244, doi:10.1186/s12936-015-0763-7.
- Ochomo, E.; Bayoh, N.M.; Kamau, L.; Atieli, F.; Vulule, J.; Ouma, C.; Ombok, M.; Njagi, K.; Soti, D.; Mathenge, E.; et al. Pyrethroid susceptibility of malaria vectors in four Districts of western Kenya. Parasites Vectors 2014, 7, 310, doi:10.1186/1756-3305-7-310.
- Kipyab, P.C.; Khaemba, B.M.; Mwangangi, J.M.; Mbogo, C.M. The physicochemical and environmental factors affecting the distribution of Anopheles merus along the Kenyan coast. Parasites Vectors 2015, 8, 221, doi:10.1186/s13071-015-0819-0.
- Mukiama, T.K.; Mwangi, R.W. Seasonal population changes and malaria transmission potential of Anopheles pharoensis and the minor anophelines in Mwea Irrigation Scheme, Kenya. Acta Trop. 1989, 4, 181–189.
- Degefa, T.; Yewhalaw, D.; Zhou, G.; Lee, M.; Atieli, H.; Githeko, A.K.; Yan, G. Indoor and outdoor malaria vector surveillance in western Kenya: Implications for better understanding of residual transmission. J. 2017, 16, 443, doi:10.1186/s12936-017-2098-z.
- Ministry of Health. The Epidemiology and Control Profile of Malaria in Kenya: Reviewing the Evidence to Guide the future Vector Control. National Malaria Control Programme, Ministry of Health. Technical support provided by the LINK Project Nairobi, Kenya, 2016. Available online: https://virtual.lshtm.ac.uk/
- wp-content/uploads/2016/11/Kenya-Epidemiological-Profile.pdf (accessed on 28 March 2019).
- Kweka, E.J.; Munga, S.; Himeidan, Y.; Githeko, A.K.; Yan, G. Assessment of mosquito larval productivity among different land use types for targeted malaria vector control in the western Kenya highlands. Parasites Vectors 2015, 8, 356, doi:10.1186/s13071-015-0968-1.
- President’s Malaria Initiative. Kenya Malaria Operational Plan FY 2019. 2019. Available online: https://www.pmi.gov/docs/default-source/default-document-library/malaria-operational-plans/fy19/fy-2019-kenya-malaria-operational-plan.pdf?sfvrsn=3 (accessed on 1 March 2019).
- Owino, E.A. Kenya needs cohesive policies and better strategies in its war against malaria in arid and semi arid areas. J. Mosq. Res. 2018, 5, 124–126.
- Bowman, D.D.; Liu, Y.; McMahan, C.S.; Nordone, S.K.; Yabsley, M.J.; Lund, R.B. Forecasting United States heartworm Dirofilaria immitis prevalence in dogs. Parasites Vectors 2016, 9, 540, doi:10.1186/s13071-016-1804-y.
- Labarthe, N.; Guerrero, J. Epidemiology of heartworm: What is happening in South America and Mexico? Parasitol. 2005, 133, 149–156, doi:10.1016/j.vetpar.2005.04.006.
- Corbet, P.S.; Williams, M.C.; Gillett, J.D. O’nyong nyong fever: An epidemic virus disease in East Africa. IV. Vector studies at epidemic sites. R. Soc. Trop. Med. Hyg. 1961, 55, 463–480.
- LaBeaud, A.D.; Banda, T.; Brichard, J.; Muchiri, E.M.; Mungai, P.L.; Mutuku, F.M.; Borland, E.; Gildengorin, G.; Pfeil, S.; Teng, C.Y.; et al. High Rates of O’Nyong Nyong and Chikungunya Virus Transmission in Coastal Kenya. PLoS Negl. Trop. Dis. 2015, 9, e0003436, doi:10.1371/journal.pntd.0003436.
- Njenga, S.M.; Mwandawiro, C.S.; Wamae, C.N.; Mukoko, D.A.; Omar, A.A.; Shimada, M.; Bockarie, M.J.; Molyneux, D.H. Sustained reduction in prevalence of lymphatic filariasis infection in spite of missed rounds of mass drug administration in an area under mosquito nets for malaria control. Parasites Vectors 2011, 4, 90, doi:10.1186/1756-3305-4-90.
- Njenga, S.M.; Kanyi, H.M.; Mutungi, F.M.; Okoyo, C.; Matendechero, H.S.; Pullan, R.L.; Halliday, K.E.; Brooker, S.J.; Wamae, C.N.; Onsongo, J.K.; et al. Assessment of lymphatic filariasis prior to re-starting mass drug administration campaigns in coastal Kenya. Parasites Vectors 2017, 10, 99, doi:10.1186/s13071-017-2044-5.
- Mwandawiro, C.S.; Fujimaki, Y.; Mitsui, Y.; Katsivo, M. Mosquito vectors of bancroftian filariasis in Kwale District, Kenya. East Afr. Med. J. 1997, 74, 288–293.
- Moraga, P.; Cano, J.; Baggaley, R.F.; Gyapong, J.O.; Njenga, S.M.; Nikolay, B.; Davies, E.; Rebollo, M.P.; Pullan, R.L.; Bockarie, M.J.; et al. Modelling the distribution and transmission intensity of lymphatic filariasis in sub-Saharan Africa prior to scaling up interventions: Integrated use of geostatistical and mathematical modelling. Parasites Vectors 2015, 8, 560, doi:10.1186/s13071-015-1166-x.
- Ogola, E.O.; Fillinger, U.; Ondiba, I.M.; Villinger, J.; Masiga, D.K.; Torto, B.; Tchouassi, D.P. Insights into malaria transmission among Anopheles funestus mosquitoes, Kenya. Parasites Vectors 2018, 11, 577, doi:10.1186/s13071-018-3171-3.
- Ogola, E.O.; Fillinger, U.; Isabella, M.V.; Jandouwe, M.; Daniel, K.; Torto, B.; Tchouassi, D.P. Malaria in Kakuma refugee camp, Turkana, Kenya: Facilitation of Anopheles arabiensis vector populations by installed water distribution and catchment systems. J. 2011, 10, 149, doi:10.1186/s13071-018-3171-3.
- Njenga, S.M.; Muita, M.; Kirigi, G.; Mbugua, J.; Mitsui, Y.; Fujimaki, Y.; Aoki, Y. Bancroftian filariasis in Kwale District, Kenya. East Afr. Med. J. 2000, 77, 245–249.
- Ciota, A.T.; Kramer, L.D. Vector-virus interactions and transmission dynamics of West Nile virus. Viruses 2013, 5, 3021–3047, doi:10.3390/v5123021.
- Burkett-Cadena, N.D.; Bingham, A.M.; Porterfield, C.; Unnasch, T.R. Innate preference or opportunism: Mosquitoes feeding on birds of prey at the Southeastern Raptor Center. Vector Ecol. 2014, 39, 21–31, doi:10.1111/j.1948-7134.2014.12066.x.
- Zittra, C.; Flechl, E.; Kothmayer, M.; Vitecek, S.; Rossiter, H.; Zechmeister, T.; Fuehrer, H.P. Ecological characterization and molecular differentiation of Culex pipiens complex taxa and Culex torrentium in eastern Austria. Parasites Vectors 2016, 9, 197, doi:10.1186/s13071-016-1495-4.
- Beji, M.; Rhim, A.; Roiz, D.; Bouattour, A. Ecophysiological characterization and molecular differentiation of Culex pipiens forms (Diptera: Culicidae) in Tunisia. Parasites Vectors 10, 3021-3047, doi:10.3390/v5123021.
- Scott, J.G.; Yoshimizu, M.H.; Kasai, S. Pyrethroid resistance in Culex pipiens Pestic. Biochem. Physiol. 2015, 120, 68–76, doi:10.1016/j.pestbp.2014.12.018.
- Centers for Disease Control. Vectors of Lymphatic Filariasis. 2018. Available online: https://www.cdc.gov/
- parasites/lymphaticfilariasis/gen_info/vectors.html (accessed on 13 March 2019).
- World Health Oganisation. Lymphatic Filariasis. 2018. Available online: https://www.who.int/news-room/fact-sheets/detail/lymphatic-filariasis (accessed on 26 March 2019).
- Turell, M.J. Members of the Culex pipiens Complex as Vectors of Viruses. Am. Mosq. Control Assoc. 2012, 28, 123–126, doi:10.2987/8756-971X-28.4.123.
- Rappole, J.H.; Hubalek, Z. Migratory birds and West Nile virus. Appl. Microbiol. 2003, 94, 47–58.
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of west nile virus. Res. Int. 2015, 2015, 376230, doi:10.1155/2015/376230.
- Kopp, A.; Gillespie, T.R.; Hobelsberger, D.; Estrada, A.; Harper, J.M.; Miller, R.A.; Eckerle, I.; Müller, M.A.; Podsiadlowski, L.; Leendertz, F.H.; et al. Provenance and geographic spread of St. Louis encephalitis virus. MBio 2013, 4, e00322-13, doi:10.1128/mBio.00322-13.
- Deardorff, E.R.; Weaver, S.C. Vector competence of Culex (Melanoconion) taeniopus for equine-virulent subtype IE strains of Venezuelan equine encephalitis virus. J. Trop. Med. Hyg. 2010, 82, 1047–1052, doi:10.4269/ajtmh.2010.09-0556.
- Samy, A.M.; Elaagip, A.H.; Kenawy, M.A.; Ayres, C.F.J.; Peterson, A.T.; Soliman, D.E. Climate change influences on the global potential distribution of the mosquito Culex quinquefasciatus, vector of West Nile virus and lymphatic filariasis. PLoS ONE 2016, 11, e0163863, doi:10.1371/journal.pone.0163863.
- Amraoui, F.; Atyame-Nten, C.; Vega-Rúa, A.; Lourenço-De-Oliveira, R.; Vazeille, M.; Failloux, A.B. Culex mosquitoes are experimentally unable to transmit zika virus. Surveill. 2016, 21, 35, doi:10.2807/1560-7917.ES.2016.21.35.30333.
- Huang, Y.S.; Ayers, V.B.; Lyons, A.C.; Unlu, I.; Alto, B.W.; Cohnstaedt, L.W.; Higgs, S.; Vanlandingham, D.L. Culex Species Mosquitoes and Zika Virus. Vector-Borne Zoonotic Dis. 2016, 16, 673–676, doi:10.1089/vbz.2016.2058.
- Phumee, A.; Chompoosri, J.; Intayot, P.; Boonserm, R.; Boonyasuppayakorn, S.; Buathong, R.; Thavara, U.; Tawatsin, A.; Joyjinda, Y.; Wacharapluesadee, S.; et al. Vertical transmission of Zika virus in Culex quinquefasciatus Say and Aedes aegypti () mosquitoes. Sci. Rep. 2019, 9, 5257, doi:10.1038/s41598-019-41727-8.
- Grossi-Soyster, E.N.; Banda, T.; Teng, C.Y.; Muchiri, E.M.; Mungai, P.L.; Mutuku, F.M.; Gildengorin, G.; Kitron, U.; King, C.H.; Desiree, L.A. Rift valley fever seroprevalence in Coastal Kenya. J. Trop. Med. Hyg. 2017, 97, 115–120, doi:10.4269/ajtmh.17-0104.
- Harbach, R. Genus Mansonia Blanchard, 1901. Mosquito Taxonomic Inventory. 2008. Available online: http://mosquito-taxonomic-inventory.info/genus-emmansoniaem-blanchard-1901 (accessed on 1 March 2019).
- Islam, M.N.; ZulKifle, M.A.; Sherwani, M.K.; Ghosh, S.K.; Tiwari, S. Prevalence of malaria, dengue and chikungunya significantly associated with mosquito breeding sites. Islam. Med. Assoc. North Am. 2011, 43, 58–67, doi:10.5915/43-2-7871.
- Torres, R.; Samudio, R.; Carrera, J.P.; Young, J.; Maârquez, R.; Hurtado, L.; Weaver, S.; Chaves, L.F.; Tesh, R.; Caâceres, L. Enzootic mosquito vector species at equine encephalitis transmission foci in the República de Panama. PLoS ONE 2017, 12, e0185491, doi:10.1371/journal.pone.0185491.
- Beranek, M.D.; Gallardo, R.; Almirón, W.R.; Contigiani, M.S. First detection of Mansonia titillans (Diptera: Culicidae) infected with St. Louis encephalitis virus (Flaviviridae: Flavivirus) and Bunyamwera serogroup (Peribunyaviridae: Orthobunyavirus) in Argentina. Vector Ecol. 2018, 43, 340–343, doi:10.1111/jvec.12320.
- Rueanghiran, C.; Lertwatcharasarakul, P.; Ruangsittichai, J. Species-specific primers for the detection of lymphatic filariasis vectors: Mansonia bonneae and Mansonia dives. Biomed. 2017, 34, 615–621.
- Ughasi, J.; Bekard, H.E.; Coulibaly, M.; Adabie-Gomez, D.; Gyapong, J.; Appawu, M.; Wilson, M.D.; David, B.; Daniel, A. Mansonia africana and Mansonia uniformis are Vectors in the transmission of Wuchereria bancrofti lymphatic filariasis in Ghana. Parasites Vectors 2012, 5, 89, doi:10.1186/1756-3305-5-89.
- Onapa, A.W.; Pedersen, E.M.; Reimert, C.M.; Simonsen, P.E. A role for Mansonia uniformis mosquitoes in the transmission of lymphatic filariasis in Uganda? Acta Trop. 2007, 101, 159–168, doi:10.1016/j.actatropica.2007.01.003.
- Epelboin, Y.; Talaga, S.; Epelboin, L.; Dusfour, I. Zika virus: An updated review of competent or naturally infected mosquitoes. PLoS Negl. Trop. Dis. 2017, 11, e0005933, doi:10.1371/journal.pntd.0005933.
- Kinyatta, N.M. Vectorial Potential of Mansonia Species in the Transmission of Wuchereria Bancrofti and Evaluation of Mosquito Collection Methods in Tana-Delta, Coastal Kenya. Master’s Thesis, Kenyatta University of Agriculture and Technology, Nairobi, Kenya, 2010.


