2. General Characteristics of the CELF Protein Family
So far, six CELF RBPs have been discovered, with a nomenclature ranging from CELF1 to CELF6. Based on their phylogenetic structure and expression pattern, these members may be divided into two subfamilies: CELF1-2 and CELF3-6. CELF1 and CELF2 are extensively expressed in numerous tissues throughout the human body, but CELF3 through 5 are mostly found in the neurological system. CELF6 is most often found in the nervous system, the kidneys, and the testes
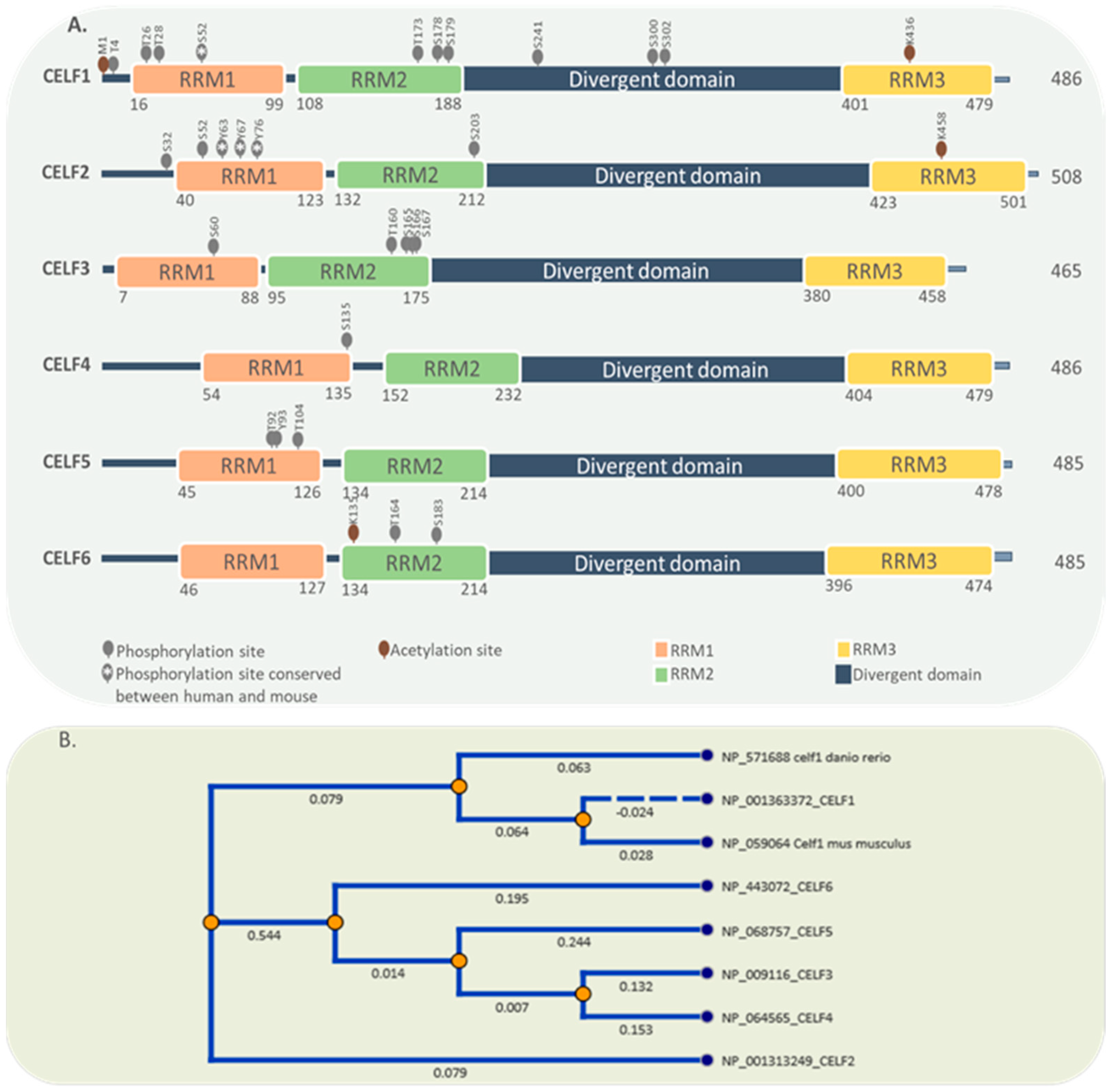
[5][11][12]. All six members have a similar structure that includes three RRMs (RNA Recognition Motifs), two in the N-terminal and one in the C-terminal, as well as a divergent linker region that can improve RBP binding affinity through conformational variations. The nuclear localization signal, which comprises a lysine/arginine-rich region in the C terminal and a region important for nuclear export in the linker domain, are two other key structures
[6][13][14]. A serine/threonine-rich phosphorylation area within the CELF1 and CELF2 linker domains could change their affinity for binding to target mRNAs
[15][16]. However, there are no reports regarding the phosphorylation of CELF2 at this site. Apart from phosphorylation, the acetylation of the lysine436 residue inside the third RRM motif has been shown to influence CELF protein–protein interactions and mRNA affinity
[17][18]. There are few studies in relation to the evolution of the CELF family members; however, their presence in xenopus, zebrafish, and mammals is clear evidence of their ontological importance and their conservation. During embryonic development CELF1 localizes in the vegetal pole of zebrafish and CELF2 in the animal pole
[19][20]. This compartmentalization suggests that each protein performs different functions and, therefore, that they must have separated very early in evolutionary terms. Future works about the evolution and function of CELF proteins would clarify how and why these proteins arose.
By generating different isoforms, the alternative splicing process modifies the structure, expression, cellular location, and functional properties of CELF family members. For example, CELF2 features three promoters that create three distinct isoforms, each of which features a unique N-terminal sequence and 5′UTR
[21]. CELF2 loses its RNA binding capacity when exon 14 is skipped; moreover, exon 9 skipping is reported for CELF4
[21][22]. Alternative splicing on 3′UTRs and C termini of CELF2 and CELF5 may affect their regulation and function
[6].
Figure 1 illustrates the structure and conservation of all six proteins of the CELF family. Reported phosphorylation and acetylation sites are obtained from PhosphoSitePlus
[23].
Figure 1. Protein domain structure and post-translational modifications of CELF family members. (A) CELF family members share the same domain structure comprising 3 RRMs and a divergent domain. Phosphorylation and acetylation sites are also shown. (B) Comparisons of the protein sequences suggest three clear branches for the evolution of CELF members. One branch for CELF1, other for CELF2 and a third for the group 3 to 6.
CELF proteins are found in the cytoplasm and nucleus, but they can also be found in stress granules, RNA neuronal granules, and on the PNC (peri-nuclear compartment) under specific circumstances
[6][24][25]. As observed most clearly in skeletal muscle loss
[26], CELF action remarkably depends on cellular location. CELF2 appears to be primarily or entirely nuclear, unlike CELF1, which is expressed in both the nucleus and the cytoplasm. As a result, despite the fact that they may bind to similar sequences, CELF2 and CELF1 are predicted to have distinct impacts on RNA processing
[7][27].
CELF RBPs are primarily involved in alternative splicing and transcript editing in the nucleus. Members of this family are all potential global alternative splicing regulators by generally recognizing CUG and UG-repeat elements
[28][29]. According to a genome-wide interaction analysis between CELF1 and its target RNAs, this RBP’s distribution at exon-intron borders, 3UTR, and even exonic regions can comprehensively affect alternative splicing events
[8][29]. However, more complex CELF1 binding sites are occasionally required for more specialized splicing processes. An alternative 3′UTR splicing site with two CELF1 binding sites separated by a long intron is required to regulate the mPGES-1 isoforms
[30]. Similarly, CELF2 has been identified as an alternative splicing factor for a high number of genes
[31]. As observed for exon 6 of LEF1 (Lymphoid Enhancer Binding Factor) and exon 2 of MKK7 (Mitogen-Activated Protein kinase 7) transcripts, CELF2 binding to a downstream or upstream region of an alternate exon can result in that exon being either included or excluded, respectively
[7]. CELF2 is additionally a regulator of 3′ UTR intron retention by competing with the U2-auxiliary factor (U2AF65) subunit. Notably, CELF2 mediates the 3′ UTR intron retention of its own transcript, as well as many others
[32]. Lastly, CELF2 has been found to be involved in another nuclear function: blocking the C to U editing of apolipoprotein B (APOB) RNA, a process that may be stimulated by CELF2 knockdown
[33].
In the cytoplasm, CELF proteins are involved in mRNA stability, translation, alternative polyadenylation (APA), and pre-miRNA maturation
[12][32][34][35][36]. CELFs regulate mRNA stability across the genome, although only specific, well-studied events are discussed in this section. CELF1 controls mRNA stability by identifying “UGUUUGUUUGU” element (GU-rich element, GRE) or GU-repeat sequences in the 3′UTR, which mechanistically appear to rely on CELF1 interaction with ribonucleases, such as poly(A)-specific ribonuclease (PARN)
[8][26]. PARN 3′-exoribonuclease destabilizes transcripts by removing poly(A) tails at the 3′ end
[37]. On the other hand, CELF2 stabilizes the cytoxygenase-2 (COX2) transcript through a different method, which involves binding to A/U-rich regions in the 3′UTR of mRNA
[5]. CELF6 has been shown to bind predominantly to certain 3′UTR regions, resulting in a substantial decrease in mRNA levels
[12].
CELF proteins have been found to regulate widespread translational activation or inhibition by binding to target transcripts’ 5′UTR or 3′UTR
[38]. For instance, the timed translation of Elavl4 (Embryonic Lethal, Abnormal Vision, Drosophila-Like 4) isoforms depends on distinct 5′UTRs controlled by CELF1 during neuronal development
[39]. Moreover, CELF1 is able to regulate P21 mRNA stability by binding to its 5′UTR upon proteasome inhibition
[40]. Binding through 3′UTR, CELF2 can inhibit the translation of MCL1 (Apoptosis Regulator, BCL2 Family Member) anti-apoptotic factor
[41]. Another example is the circularization of SHMT1 (Serine Hydroxymethyltransferase 1) mRNA and its internal ribosome entry site (IRES)- mediated translation, which is provided with CELF1-hnRNPH (Heterogeneous Nuclear Ribonucleoprotein H1) proteins, implying a 5′/3′ interaction in the SHMT1 transcript
[42]. Interestingly, following CELF2 depletion, hnRNPC (Heterogeneous Nuclear Ribonucleoprotein C) translation efficiency is shown to be reduced. This is mostly due to the hnRNPC mRNA’s lower translation elongation efficiency
[34]. Further pathway study indicates that the knockdown of the CELF1 decreases the expression of some ribosomal proteins in the 40S and 60S subunits, indicating a broader function of the CELF1 in translation
[29].
CELF proteins are known to regulate alternative polyadenylation (APA) via two mechanisms. Firstly, CELF1 has been shown to bind to c-FOS (Fos Proto-Oncogene, AP-1 Transcription Factor Subunit) and TNFα (Tumor Necrosis Factor-Alpha) transcripts in a HeLa cell extract and promote deadenylation by interacting with PARN, indicating that CELF1 helps both deadenylation and mRNA stability
[35]. Secondly, CELF2 prevents polyadenylation machinery from binding to RNA by competing with CFI (cleavage factor I) and CstF (cleavage stimulatory factor) components in a cell signal-dependent way
[32].
Recently, CELF2 was reported as a potential regulator of pre-miR155 maturation. CELF2 is reported to be associated with IL10 (Interleukin-10) to inhibit the maturation of pre-miR155 by blocking KSRP (KH-Type Splicing Regulatory Protein). In this way, it can be considered as negative regulator of miR155
[36]. Similar results are reported for miR140, where CELF1/2 binding interferes with DICER binding to miR140 and thus inhibits its maturation
[43].
Finally, CELF RBPs are present in stress granules, neuronal RNA granules, and the peri-nucleolar compartment (PNC). Stress granules (SGs) are dynamic tiny ribonucleoprotein condensates playing functional roles in mRNA processing and degradation and are found in all eukaryotes during cell stress conditions
[44]. CELF1 and CELF2 have been found to be required for the trafficking, stability, and translation of p21/CDKN1A and COX-2 to SGs respectively
[40][45]. Additionally, CELF4 associates with very large neuronal RNA granules to mediate neurotransmission by regulating the stability, translation, and/or localization of many downstream target mRNAs
[25].
PNCs are specialized subnuclear organelles located adjacent to the nucleolus. They are associated with various solid malignant tumors. This region contains transcripts produced by RNA polymerase III, small nuclear RNAs, and many RBPs, including CELF1
[46][47]. In PNCs, CELF1 associates with the RNA component of RNAse MRP (mitochondrial RNA processing) and PTBP (polypyrimidine tract binding protein), interacting with newly synthesized RNA. However, CELF1 enrichment in PNCs may restrict it from performing its cytoplasmic functions
[6][48].
In consequence, CELF members are highly divergent, pleiotropic RBPs that are responsible for global alterations in numerous cell types and disease situations
[5]. However, CELF proteins are not the only multifunctional RBPs. In two previously published studies, various RBPs influencing different cancers were described
[49][50]. The principal functions of CELF family proteins are depicted in
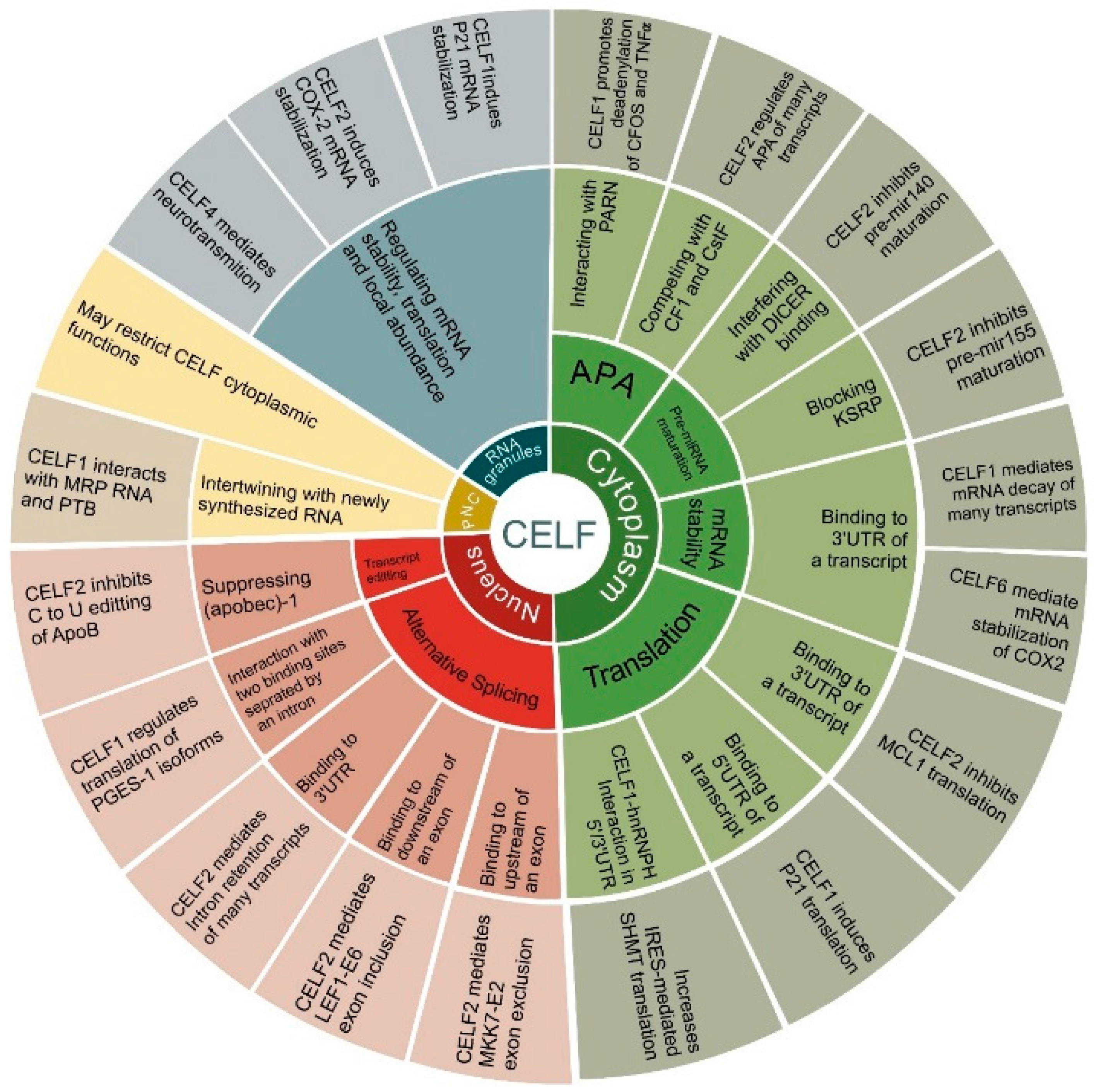
Figure 2. Furthermore, they are controlled by numerous biological elements that are discussed specifically in relation to cancer in this article.
Figure 2. Cellular functions of CELF family members. The circular diagram is divided into 4 main branches illustrating CELF subcellular locations, including the cytoplasm, the nucleus, RNA granules and the peri-nuclear compartment (PNC). Next, each branch is divided into CELFs’ main functions, including translation, mRNA stability, pre-miRNA maturation and alternative polyadenylation (APA) in the cytoplasm, alternative splicing and transcript editing in the nucleus, regulating mRNA stability, translation, local abundance in RNA granules, and, finally, intertwining with newly synthesized RNA in PNCs. In each case, the main mechanism and some representative instances are included for each function. These are discussed in depth in the text.