1000/1000
Hot
Most Recent
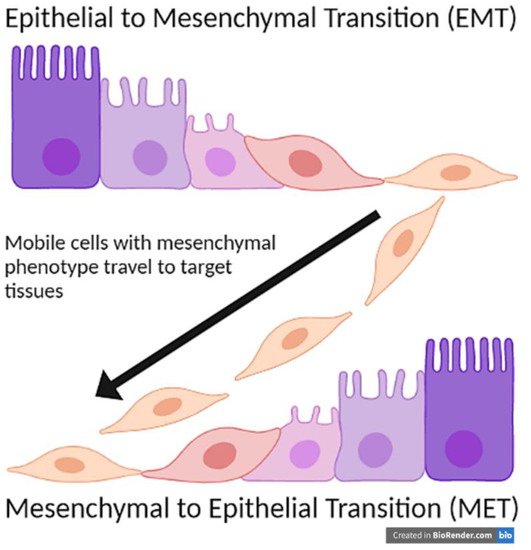
Cripto-1 is an essential protein for human development that plays a key role in the early phase of gastrulation in the differentiation of an embryo as well as assists with wound healing processes. Importantly, Cripto-1 induces epithelial to mesenchymal transition to turn fixed epithelial cells into a more mobile mesenchymal phenotype through the downregulation of epithelial adhesion molecules such as E-cadherin, occludins, and claudins, and the upregulation of mesenchymal, mobile proteins, such as N-cadherin, Snail, and Slug. Consequently, Cripto-1’s role in inducing EMT to promote cell motility is beneficial in embryogenesis, but detrimental in the formation, progression and metastasis of malignant tumors. Indeed, Cripto-1 is found to be upregulated in most cancers, such as breast, lung, gastrointestinal, hepatic, renal, cervical, ovarian, prostate, and skin cancers. Through its role in EMT, Cripto-1 can remodel cancer cells to enable them to travel through the extracellular matrix as well as blood and lymphatic vessels to metastasize to different organs. Additionally, Cripto-1 promotes the survival of cancer stem cells, which can lead to relapse in cancer patients.