Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 + credit
+ credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Emma J. Murphy | + 4501 word(s) | 4501 | 2021-08-19 04:22:32 | | | |
| 2 | Lindsay Dong | -5 word(s) | 4496 | 2021-08-19 08:43:01 | | | | |
| 3 | Lindsay Dong | -4037 word(s) | 459 | 2022-04-13 12:26:16 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Murphy, E.; Rezoagli, E.; Major, I.; Rowan, N.; Laffey, J. β-Glucans. Encyclopedia. Available online: https://encyclopedia.pub/entry/13331 (accessed on 07 February 2026).
Murphy E, Rezoagli E, Major I, Rowan N, Laffey J. β-Glucans. Encyclopedia. Available at: https://encyclopedia.pub/entry/13331. Accessed February 07, 2026.
Murphy, Emma, Emanuele Rezoagli, Ian Major, Neil Rowan, John Laffey. "β-Glucans" Encyclopedia, https://encyclopedia.pub/entry/13331 (accessed February 07, 2026).
Murphy, E., Rezoagli, E., Major, I., Rowan, N., & Laffey, J. (2021, August 19). β-Glucans. In Encyclopedia. https://encyclopedia.pub/entry/13331
Murphy, Emma, et al. "β-Glucans." Encyclopedia. Web. 19 August, 2021.
Copy Citation
β-glucans are complex polysaccharides that are found in several plants and foods, including mushrooms. β-glucans display an array of potentially therapeutic properties.
β-glucan
clinical trials
biomedicine
immunomodulation
metabolism
β-glucans/Beta-glucans are a large class of complex polysaccharides that can be found in an abundance of sources. Depending on origin, β-glucans can be classified as cereal or non-cereal derived. Cereal sources of β-glucans include oat and barley and non-cereal sources can include mushroom, algae, bacteria and seaweed [1]. β-glucans are biologically active compounds that have been widely reported to improve health [2]. Specific to this group of polysaccharides is a 1,3 beta-glycosidic linked backbone; separate to this, the polysaccharide can take many forms, dictated by origin.
There is a growing interest in foods that have the potential to lower the risk or incidences of chronic diseases or promote lifespan as well as have anti-aging properties. This has led to an increase in awareness of the effect of diet on health [3][4].
In 1979, Stephen DeFelice devised the term nutraceutical, which may be isolated nutrients, dietary supplements, genetically engineered foods and herbal products [5]. Nutraceuticals are defined as a food or food component that provides medical or health benefits, including prevention and/or treatment of disease [5]. Similarly, bioactive compounds are defined as “essential and non-essential compounds that occur in nature, are part of the food chain and are shown to have an effect on human health” [6].
Bioactive substances in food provide health benefits beyond the nutritional benefits of the product [7]. β-glucans are reported to be both a bioactive and a nutraceutical. Their therapeutic effects can also be largely classified into two categories, metabolic/GI effects or immune-modulatory effects, which is largely based on structure, determined by source [1][8].
Metabolic effects are usually observed with cereal derived β-glucans. Effects include modulation of the gut microbiome, cholesterol reduction and decreased cardiovascular and diabetic risk. Non-cereal β-glucans are associated with immune-modulatory effects, anti-tumor effects, wound healing and alleviation of immune-related conditions, as demonstrated in Figure 1 [1]. β-glucans are also administered as an animal and fish feed additive to increase [9][10]. These molecules also have applications in the food industry for thickening and for gelation purposes [11].
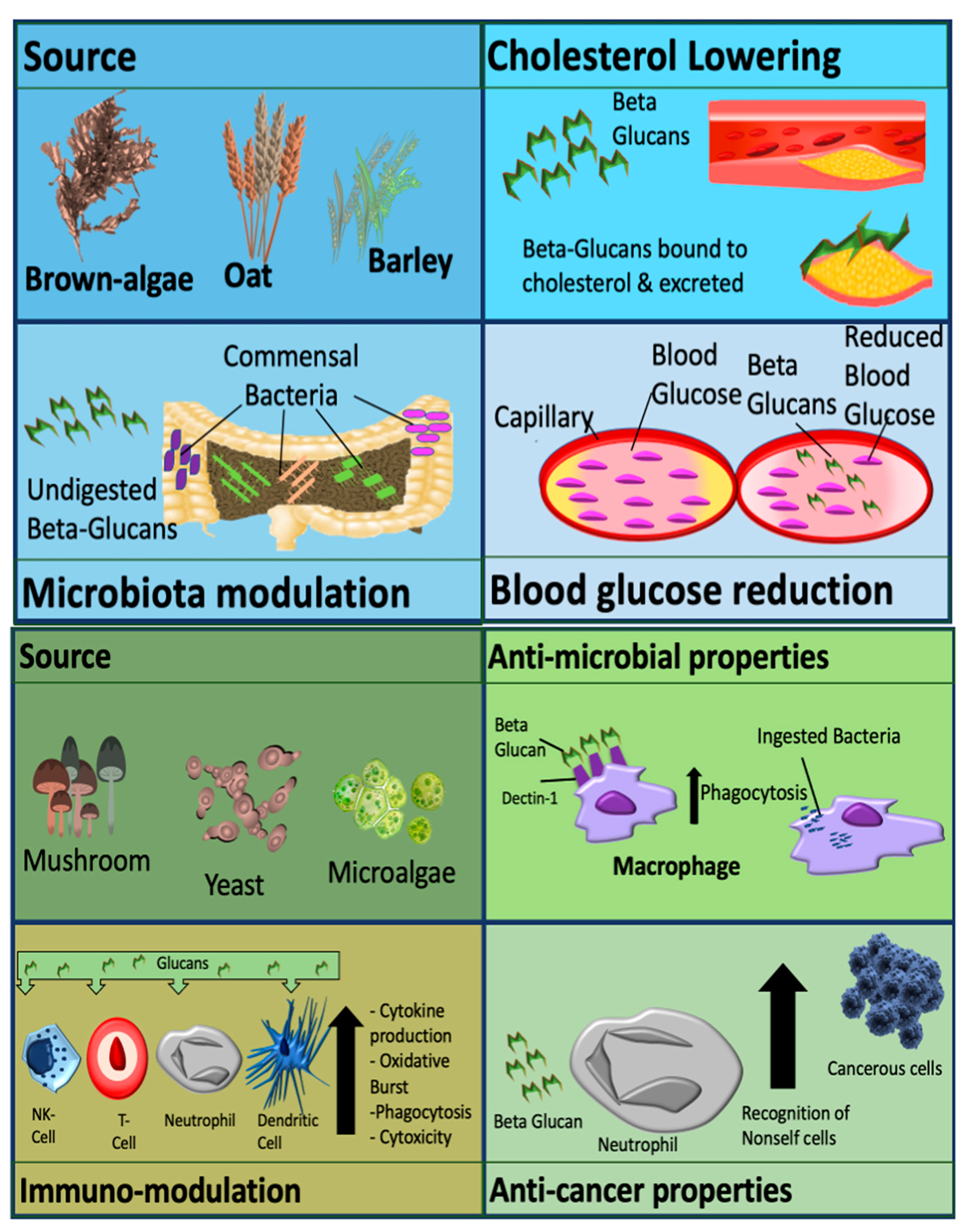
Figure 1. Mechanisms and activity of β-glucan which are dependent on source. β-glucan can be classified as cereal derived (upper panel), or non-cereal derived (lower panel). Picture originally published in [1]. Modified with permission.
In this encyclopedia entry on β-glucans, we start from their initial discovery, then examine β-glucan sources, characterize their complex and diverse structures, and examine the implications of their structural variations on their activity profile. The therapeutic potential in different disease conditions is then discussed, and the barriers to fully realizing this potential is dissected in some detail. Finally, we examine other uses of β-glucan in animal health and their application in the food industry.
References
- Murphy, E.J.; Rezoagli, E.; Major, I.; Rowan, N.J.; Laffey, J.G. β-Glucan Metabolic and Immunomodulatory Properties and Potential for Clinical Application. J. Fungi 2020, 6, 356.
- Wang, Q.; Sheng, X.; Shi, A.; Hu, H.; Yang, Y.; Liu, L.; Fei, L.; Liu, H. β-Glucans: Relationships between modification, conformation and functional activities. Molecules 2017, 22, 257.
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100.
- Ahnen, R.T.; Jonnalagadda, S.S.; Slavin, J.L. Role of plant protein in nutrition, wellness, and health. Nutr. Rev. 2019, 77, 735–747.
- Kalra, E.K. Nutraceutical-Definition and introduction. AAPS PharmSci 2003, 5, 27.
- Frank, J.; Fukagawa, N.K.; Bilia, A.R.; Johnson, E.J.; Kwon, O.; Prakash, V.; Miyazawa, T.; Clifford, M.N.; Kay, C.D.; Crozier, A.; et al. Terms and nomenclature used for plant-derived components in nutrition and related research: Efforts toward harmonization. Nutr. Rev. 2020, 78, 451–458.
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-Martins, M. Methods for determining bioavailability and bioaccessibility of bioactive compounds and nutrients. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Elsevier: Amsterdam, The Netherlands, 2019; pp. 23–54.
- Ulmius, M.; Önning, G.; Nilsson, L. Solution behavior of barley β-glucan as studied with asymmetrical flow field-flow fractionation. Food Hydrocoll. 2012, 26, 175–180.
- Rodrigues, M.V.; Zanuzzo, F.S.; Koch, J.F.A.; de Oliveira, C.A.F.; Sima, P.; Vetvicka, V. Development of Fish Immunity and the Role of β-Glucan in Immune Responses. Molecules 2020, 25, 5378.
- Byrne, K.A.; Loving, C.L.; McGill, J.L. Innate Immunomodulation in Food Animals: Evidence for Trained Immunity? Front. Immunol. 2020, 11.
- Ahmad, A.; Anjum, F.M.; Zahoor, T.; Nawaz, H.; Dilshad, S.M.R. Beta glucan: A valuable functional ingredient in foods. Crit. Rev. Food Sci. Nutr. 2012, 52, 201–212.
More
Information
Subjects:
Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.8K
Online Date:
19 Aug 2021
Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No