2. Nanoparticles Characterization and XTT Analysis
Size is a fundamental factor in determining the biological activity of nanoparticles. In general, it has been observed that the smaller the size, the higher the activity
[18]. Nevertheless, very small particles can exert toxic effects on cells: sometimes nanoparticles below 10 nm are not suitable for biological experiments because they can, for instance, cause hemolysis
[19]. Nevertheless, the presence of capping on their surface can easily modulate their activity, making them less toxic and more active and biocompatible. Therefore, it is important to find a balance between activity and toxicity by tuning the dimensions of the nanoparticles. A good way to do this is to control the pH of the solution in which the nanoparticles are synthesized, knowing that in general higher pH corresponds to smaller particles
[20]. Consequently, a procedure already in use was modified to control the dimension of the AgNPs by tuning the pH in solution
[21]. Hence, three different batches of
Artemisia–AgNPs have been prepared at distinct pH values (7, 8, and 9) in order to examine whether, as expected, the pH change was able to modify their size, but also to influence their capping and, therefore, their biological behavior. Ultracentrifugation at 13,000 rpm was introduced to increase the yield of the recovered AgNPs and to shorten the workout procedure, significantly improving the synthetic process.
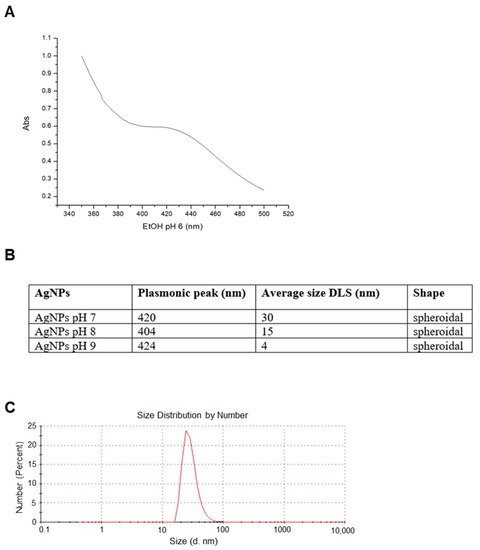
A full characterization of the AgNPs was carried out in order to study their structural features. UV-vis spectra showed a plasmonic peak around 420 (
Figure 1A), suggesting a spheroidal shape for these nanoparticles with a probable size between 10 and 30 nm
[22]. DLS analysis showed that the size distribution of AgNPs changed according to the pH of the reaction solution. As expected, an increase in the pH determined a decrease in the hydrodynamic radius
[23], which turned out to be around 30 nm for AgNPs at pH 7 (
Figure 1B), around 15 nm for AgNPs at pH 8, and around 4 nm for AgNPs at pH 9. Measurements taken after weeks from their synthesis evidenced that the dimensions remained constant over time, indicating that these particles are rather stable.
Figure 1. Artemia-AgNPs characterization. (A) Table indicating the major properties of all three nanoparticles. (B) Selected UV-vis spectrum for AgNPs at pH 7 showing the plasmonic peak at 420 nm. (C) Selected DLS measurements for AgNPs at pH 7.
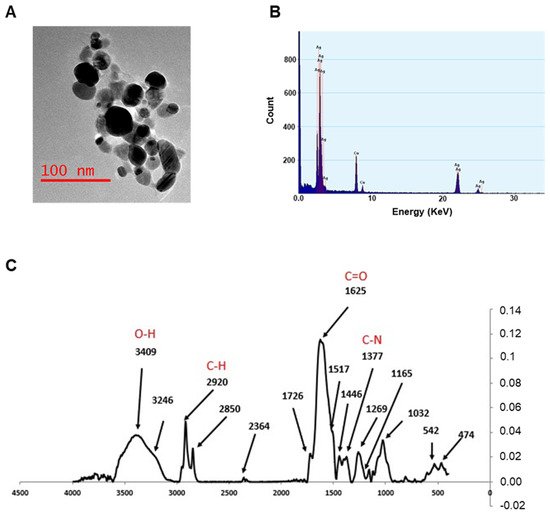
TEM analysis confirms the information on the size and morphology of the AgNPs, which were, in almost all cases, spheroidal in shape, with average dimensions well below 50 nm. The TEM image shown in Figure 2A displays the morphology of the nanoparticles obtained at pH 7, characterized by a spheroidal shape and no aggregation in water. EDX profile was used to evaluate the elemental structure of the nanoparticles (Figure 2B), underlining their silver composition. The other elements presented in the spectra are due to the holey carbon/copper grid used.
Figure 2. TEM analysis, EDX profile, and FTIR measurements. (A) TEM image for AgNPs at pH 7. (B) EDX profile showing the presence of elemental Ag (Cu signals are due to the grid used). (C) FTIR spectra of AgNPs synthesized at pH 7. Spectra were acquired in the range 400–4000 cm−1.
The FTIR measurements were performed by recording the signals in the range of 400–4000 cm−1 with a resolution of 4 cm−1, in order to acquire more information on the capping molecules. FTIR spectra from all samples were almost identical, indicating an analogous composition (Figure 2C). The bands observed suggest that amines, proteins, or (poly)phenolic compounds of Artemisia leaf extracts may be involved in the formation of the capping. A broad band from 3100 to 3600 cm−1 incorporates both OH and NH stretching frequencies, while the signals around 2900 cm−1 can be attributed to the sp3 CH stretching mode of hydrocarbon moieties, and those between 1630 (νC=O) and 1000 cm−1 are most probably due to amidic compounds (i.e., proteins and enzymes) or amines.
The characterization of the three batches of AgNPs showed that, while they were all spheroidal in shape and shared the same capping molecules, the pH at which they were obtained was actually able to influence their dimensions, with a higher pH corresponding with smaller nanoparticles as expected.
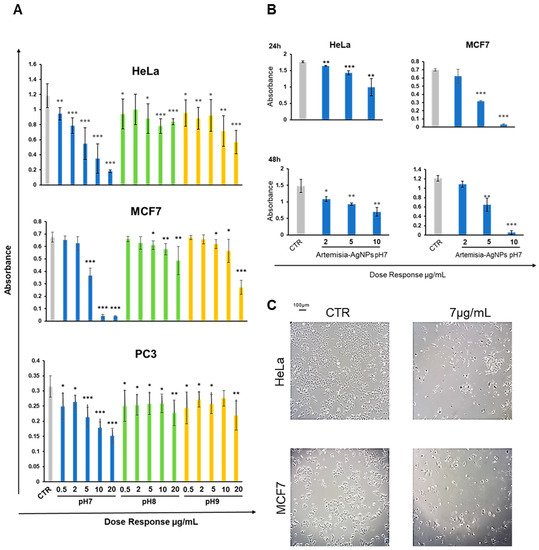
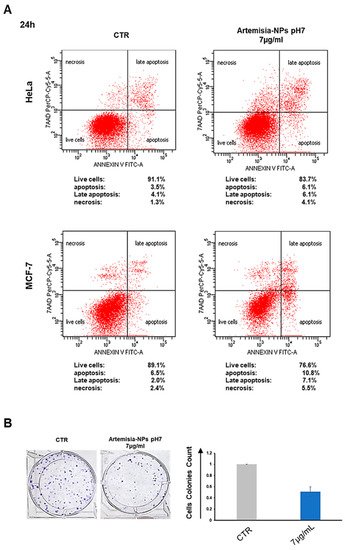
To check the antitumor effects of our AgNPs, we treated different cell cancer lines (HeLa, MCF-7 and PC3) with increasing doses (from 0.5 to 20 µg/mL) of Artemisia–AgNPs (pH 7, pH 8 and pH 9) in order to evaluate cell viability by XTT assay. We observed a dose-dependent anti-proliferative effect of Artemisia–AgNPs. The decrease in viability was more relevant for the Artemisia–AgNPs pH7 compared to the others, with a decrease in cell viability in a dose dependent manner (
Figure 3A). Other studies demonstrated the ability of AgNPs to inhibit cell proliferation, inducing cell death
[24][25]. Moreover, it was shown that the cytotoxic action is influenced by size and surface area of AgNPs
[26]. Regarding the distinct action of AgNPs due to the different sizes, recent evidence suggests that the appropriate size for cellular uptake, in order to achieve the greatest biological activity of nanoparticles without the risk of hemolysis, is around 40–50 nm
[27].
Figure 3. XTT assay. (A) HeLa, MCF-7, and PC3 were treated with increasing doses (0.5–20 µg/mL) of each Artemisia–AgNPs (pH 7, pH 8, and pH 9). An XTT assay was performed to evaluate cell viability. (B) XTT assay of Artemisia–AgNPs pH7 on HeLa and MCF-7. HeLa and MCF-7 were cultured with 2, 5, and 10 µg/mL of Artemisia–AgNPs pH 7. Cell viability was evaluated using XTT assay. (C) Microscope images. HeLa and MCF-7 treated for 24 h with 7 µg/mL of Artemisia–AgNPs pH 7. Data were analyzed using Student’s t test, * = p value < 0.05, ** = p value < 0.01, *** = p value < 0.001.
Moreover, as reported in
Figure 3A, the anti-proliferative activity of Artemisia–AgNPs pH7 is more evident on HeLa and MCF-7 compared to PC3 (
Figure 3A). In accordance with our results, Sadegh et al. observed that AgNPs synthesized using green synthesis showed more selective activity against an MCF-7 cell line
[28].
Based on these data, we selected Artemisia–AgNPs pH7 for further analysis in order to clarify their specific effects on HeLa and MCF-7. Therefore, to validate the cytotoxic effects, XTT assay was repeated at 24 h and 48 h, using concentrations of 2, 5, and 10 µg/mL of Artemisia–AgNPs pH 7. As showed in Figure 3B, Artemisia–AgNPs pH7 revealed a high anti-proliferative effect, especially at the concentration between 5 and 10 µg/mL. Thereby, for the following experiments, we considered 7 µg/mL of Artemisia–AgNPs pH7 to be the ideal concentration to significantly reduce cell viability. The microscope images in Figure 3C clearly show the cells after 24 h of treatment with 7 µg/mL of Artemisia–AgNPs pH7. Compared to the control, most cells suffered when detached from the plate and died after treatment.
3. Cell Cycle Impact
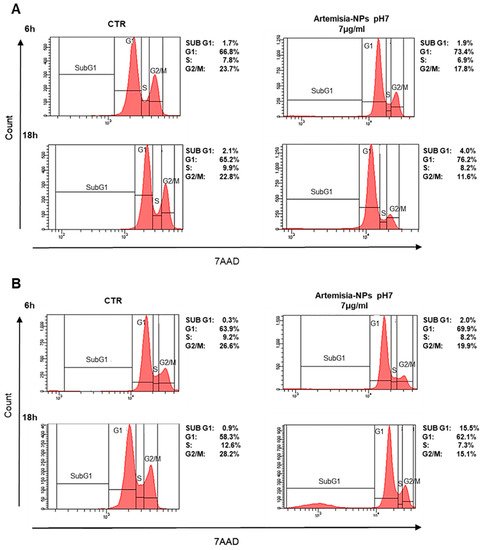
In order to evaluate the impact on the cell cycle, cells were treated with 7 µg/mL of Artemisia–AgNPs for 6 h and 18 h. Interestingly, a significant reduction in the number of HeLa cells in G2/M phase was found after only 6 h of treatment in comparison to control samples (
Figure 4A). At 18 h of incubation, the arrest of cells in the G0/G1 phase was evident (
Figure 4A). Similar results were found in MCF-7 cell lines treated with Artemisia–AgNPs (
Figure 4B). Especially, after 18 h of incubation, the percentage of cells in the G2/M phase strongly decreased in association with an increase in G0/G1 phase. Moreover, we observed a relevant increment of the SubG1 phase, an index of cellular death (
Figure 4B). The impact on cell cycle of cancer cells mediated by AgNPs was analyzed by other studies. The exposition to 10–50 μg/mL of AgNPs for 24 h determined a relevant increase in the SubG1 population on HeLa cells and cells were not able to go through G2 checkpoint
[29]. In contrast to our results, other authors studied AgNPs on lung epithelial cells and glioblastoma cells, observing an increase in G2/M-phase cells accompanied by a decrease in the G1 population after treatment
[30]. This divergence in cell cycle data may be due to different factors including cell lines, distinct synthesis of AgNPs, and presence of capping able to change the biological behaviors of AgNPs. Nonetheless, further investigations are needed to better understand the cell cycle effects mediated by AgNPs.
Figure 4. Cell cycle analysis. (A) HeLa cells were treated with 7 µg/mL of Artemisia–AgNPs pH 7 or left untreated. The cell cycle was evaluated after 6 h and 18 h by flow cytometry. (B) MCF-7 cells were treated to the same condition as HeLa and cell cycle effects were evaluated by flow cytometry.
5. Artemisia–AgNPs Impact on Gene Expression
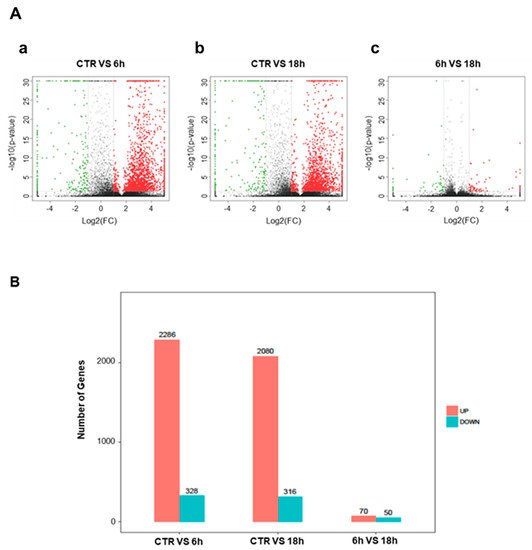
To elucidate the mechanism of action of Artemisia–AgNPs, we performed RNA-sequencing on HeLa cells in order to have a wide overview of the impact on gene expression. After 6 h and 18 h of treatment, we observed several differentially expressed genes (DEGs), using parameter the log2 fold-change values and p-values as reference indicators for significant difference (Figure 6A). Figure 6B shows in detail the gene variations: 2286 up-regulated and 328 down-regulated genes after 6 h of treatment, while after 18 h of treatment 2080 up-regulated and 316 down-regulated genes are shown.
Figure 6. Differentially expressed genes. (A) Volcanic map of differential genes. The DEGs set after 6 h and 18 h of Artemisia–AgNPs treatment. Red dots indicate up-regulated genes, green dots represent down-regulated genes, and black dots indicate genes with no significant difference. (|logFC| ≥ 2 and p value < 0.05). a: CTR vs. 6 h; b: CTR vs. 18 h; c: 6 h vs. 18 h. (B) Histogram of differential genes. The DEGs set disturbed by Artemisia–AgNPs treatment (|logFC| ≥ 2 and p value < 0.05).
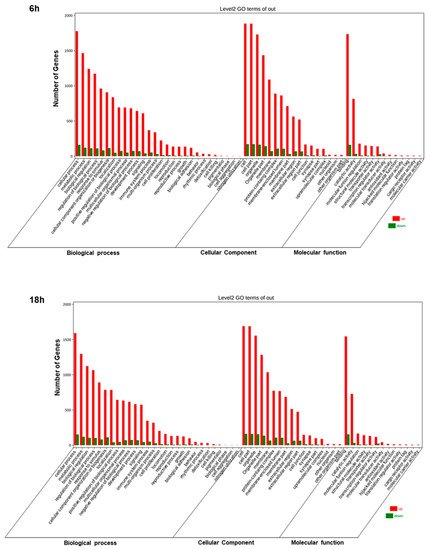
Afterward, we employed OmicShare tools, an online software, for the gene ontology (GO) analysis, enabling the study of the functions of the differentially expressed genes (DEGs). In detail, through these tools it is possible to evaluate the cellular components, biological processes, and molecular functions correlated with DEGs. We found similar results at 6 h and 18 h. Artemisia–AgNPs appear to work at an intracellular level, with enriched genes mainly expressed in organelles and their membranes (Figure 7). Regarding the biological processes, the main up-regulated genes were correlated with metabolic processes and biological regulation. The molecular functions of DEGs were mainly involved in binding, catalytic activity, and molecular function regulator.
Figure 7. GO analysis. GO annotation for 6 h and 18 h of treatments of DEGs. The number of enriched genes is presented.
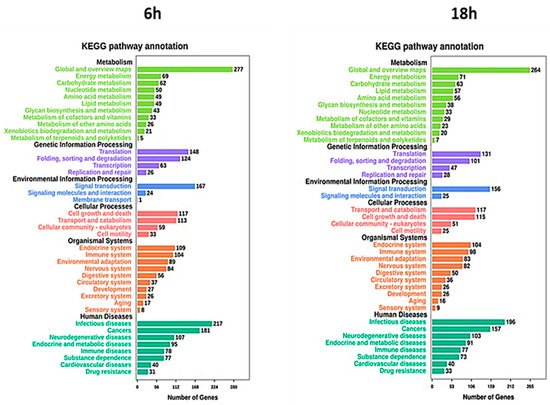
To further characterize the differentially expressed genes, investigating the metabolic and signal transduction pathways involved, KEGG pathway analysis was performed. The analysis showed a strong association between DEGs and cellular metabolism, translation, and folding processes (Figure 8). Moreover, several DEGs were also correlated with signal transduction and cell growth and death. Regarding human diseases, the DEGs correlated with Artemisia–AgNPs treatment are mainly enriched in infections and cancers.
Figure 8. KEGG analysis. The pathways involved in the DEGs were analyzed using KEGG analysis. The number of enriched genes and pathway terms are presented.
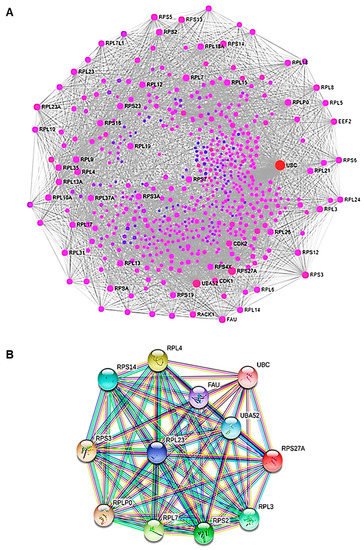
To investigate the protein–protein interaction (PPI) network, we used an online mapping tool (NetworkAnalyst3.0) (
Figure 9A). The PPI network analysis led to the evaluation of the top 15 genes, classified as hub genes by degree. The following are the names of the hub genes: UBC, UBA52 (over-expressed during hepatoma cell apoptosis), RPS27A (cell malignant transformation), RPS3 (cell apoptosis regulation), FAU (down-regulated in human prostate, breast, and ovarian cancers), RPL7 (cell apoptosis regulation), RPL23A, RPL4 (self-translation regulation—
E. coli), RPLP0 (tumor progression, invasion, metastasis), RPS5, RPL9, RPS14, RPS2, RPL3, and RPL23. These genes encode for ubiquitin-ribosomal proteins, which are involved in a variety of biological events, including cellular mechanisms correlated with cancer formation and progression. Ubiquitin is a highly conserved regulatory protein that covalently binds proteins, leading to post-translational modification of proteins. Ubiquitination is a crucial event for cell cycle progression and cell proliferation. Alterations of this process are associated with cancer development
[34]. In order to clarify the association between the hub genes, we also performed a PPI analysis of the hub genes (
Figure 9B). Therefore, through the functional and pathway enrichment analysis, we identified the involvement of these 15 hub genes in several biological processes, including regulation of gene expression, DNA damage response, genome nucleotide-excision repair, and in ribosomes (
Table 1).
Figure 9. PPI analysis. (A) The PPI network of DEGs. The nodes represent the differential genes and the connections between the nodes reveal the interactions between the genes. The size of the nodes indicates the degree of the genes, and the different colors represent the distinct gene expression (red are up-regulated genes and violet are less up-regulated). (B) The PPI network of hub genes. Co-expression network with top hub DEGs.
Table 1. The top 15 hub genes enrichment with GO and KEGG.
| Category |
Term |
Description |
Gene Count |
p Value |
| Biological Process (GO) |
GO:0010629 |
Negative regulation of gene expression |
12 |
2.33 × 10−12 |
| Biological Process (GO) |
GO:0090304 |
Nucleic acid metabolic process |
12 |
5.05 × 10−8 |
| Biological Process (GO) |
GO:00427969 |
DNA demage response, detection of DNA demage |
4 |
9.62 × 10−8 |
| Biological Process (GO) |
GO:0042276 |
Error-prone translesion synthesis |
4 |
2.46 × 10−6 |
| Biological Process (GO) |
GO:0006297 |
Nucleotide-excision repair, DNA gap filling |
3 |
3.52 × 10−6 |
| Biological Process (GO) |
GO:0070911 |
Global genome nucleotide-excision repair |
3 |
4.76 × 10−6 |
| Biological Process (GO) |
GO:0000122 |
Negative regulation of transcription by RNA polymerase II |
5 |
1.05 × 10−3 |
| KEGG_PATHWAY |
Hsa03010 |
Ribosome |
11 |
2.96 × 10−20 |
 +1 credit
+1 credit