 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | David Ramiro-Cortijo | + 3724 word(s) | 3724 | 2021-06-15 09:05:34 | | | |
| 2 | Conner Chen | Meta information modification | 3724 | 2021-06-22 05:23:53 | | |
Video Upload Options
Cardiovascular disease remains a leading cause of mortality worldwide. Unresolved inflammation plays a critical role in cardiovascular diseases development. Specialized Pro-Resolving Mediators (SPMs), derived from long chain polyunsaturated fatty acids (LCPUFAs), enhances the host defense, by resolving the inflammation and tissue repair. In addition, SPMs also have anti-inflammatory properties. These physiological effects depend on the availability of LCPUFAs precursors and cellular metabolic balance. It would be interesting to know the role that SPMs play in relation to metabolic variables in neonatal and adult cardiovascular physiology. This could open the door to new therapeutic and nutritional intervention strategies, both in adults and neonates.
1. Background
Despite encouraging advancement in the field of public health, cardiology, and scientific discovery for the prevention and treatment, cardiovascular diseases are a leading cause of mortality and are one of the major public health concerns globally. Cardiovascular diseases are responsible for 30% of deaths worldwide [1][2][3]. Even after surviving myocardial infarction or stroke, the likelihood of developing secondary complications is high, and have significant health and economic burden through hospitalizations and follow-up clinical care.
In addition to lifestyle, nutrition also plays an important role in the etiology and treatment of cardiovascular diseases. Nutrient-based approaches have been suggested to reduce the risk of developing cardiovascular diseases by reducing various risk factors such as hyperlipidemia, diabetes, metabolic syndrome, overweight/obesity, and inflammatory mediators [4][5]. The cardioprotective effect of diet could be associated with an improved lipid profile, decreased blood pressure, antioxidant properties, and decreased platelet activation. It is now widely accepted that chronic inflammation plays a crucial role in the development of cardiovascular diseases. During infection and injury, innate immune cells are recruited to the site of tissue damage and produce classical eicosanoids that are highly pro-inflammatory. However, excessive pro-inflammatory response can cause cell damage and occasionally apoptosis/necrosis. Failure of macrophages to clear apoptotic cells prolongs the inflammation [6]. Resolution of inflammation is regulated by a class of bioactive lipid mediators, called specialized pro-resolving mediators. Impaired resolution of inflammation results in chronic inflammation that is associated with cancers, autoimmune diseases, metabolic, cardiovascular, and neurodegenerative diseases resulting in organ dysfunction [6][7]. Acute and chronic inflammation coexist over long periods. Uncontrolled inflammation and failure to resolve the inflammation is the underpinning of several prevalent human diseases, including cardiovascular diseases. Furthermore, finding new ways to target the inflammatory response is gaining attraction as a new therapeutic approach to treat these diseases [8].
Lipids are an important structural component of the cell membrane and the susceptibility of lipid membranes to oxidative damage is dependent on the degree of unsaturation of its fatty acids. Evidence suggests that dietary fatty acids, more specifically long-chain polyunsaturated fatty acids (LCPUFAs), are protective against cardiovascular diseases [9] and these beneficial effects are mediated by LCPUFAs derived lipid mediators [10][11]. LCPUFAs serve as precursors for eicosanoid production (prostaglandins, prostacyclins, thromboxanes, and leukotrienes), which are important regulators of thrombocyte aggregation, inflammatory response, leukocyte functions, vasoconstriction, vasodilatation, blood pressure, and bronchial constriction.
2. The Role of Fatty Acids in Cardiovascular Development and Physiology
The perinatal life is a critical period in the development and physiology of cardiac tissue. Higher cardiac energy metabolism is required in response to early postnatal oxygen levels, and increased cell proliferation is needed to attain normal neonatal growth and development [12][13][14]. Cardiomyocyte regeneration is a highly energy-consuming process, and changes in energy metabolism happen as the neonatal cardiac development evolves rapidly within a short postnatal period; afterward, the cardiomyocytes exit the cell cycle. Fetal cardiomyocytes use glycolysis as the main source of energy during proliferation [15][16]. During early postnatal development, there is a shift from glycolysis to using fatty acid oxidation (FAO) [17][18], possibly due to increased energy demand during the transitioning from a fetal to postnatal period [19], suggesting a correlation between cardiomyocyte proliferation and high oxidative energy metabolism. It is not clear whether this heart metabolic shift accounts for cardiomyocyte proliferation and hypertrophy or it is a consequence of increased oxygen availability in postnatal environment. Animal studies have shown that in younger animals, FAO facilitates cardiomyocyte proliferation and hypertrophic growth [16]. Altered FAO could be associated with cardiac diseases [20][21]. Increased levels of oxygen during transition, from fetal to postnatal period, results in significantly increased oxidative stress as a result of the production of reactive oxygen species (ROS) with subsequent damage to cardiac tissue. These data could suggest that reducing the oxygen level and increased antioxidant levels might be useful to maintain the proliferative capacity of neonatal cardiomyocytes for a longer time [12][22]. These data suggest that the antioxidant could be used as a relevant therapy to extend the critical time window for cardiomyocyte proliferation by upregulating mitochondrial biogenesis and decrease mitophagy [12][23].
Cardiolipin (CL) is a phospholipid that is exclusively present in the inner mitochondrial membrane, where it is essential for optimal functions of various key enzymes involved in mitochondrial respiration [24][25]. Linoleic acid is one of the major fatty acids of CL in human myocardium [26]. Evidence has shown that CL must contain four linoleic acid side chains for optimal mitochondrial function, and loss of linoleic acid content in CL resulted in human and rat models of heart failure [26]. Furthermore, diet supplementation containing high linoleic acid attenuated contractile failure by improving mitochondrial dysfunction in rat models of hypertensive heart failure [27].
2.1. LCPUFAs and Cardiovascular Diseases
LCPUFAs are a group of essential fatty acids that can classify into two main categories: omega-3 (n-3) and omega-6 (n-6), depending on the position of the first double bond from the methyl end group of the fatty acid. Linoleic acid (LA; 18:2n-6) and α-linolenic acid (ALA; 18:3n-3) are the precursors of n-3 and n-6 LCPUFAs, respectively. The metabolic pathway involves a series of desaturase and elongase enzymes leading to the production of eicosapentaenoic acid (EPA; 22:5n-3) from ALA or arachidonic acid (ARA; 20:4n-6) from LA, based on the availability. EPA is then elongated and desaturated to produce docosahexaenoic acid (DHA; 22:6n-3).
Human studies have shown limited endogenous synthesis of DHA from n-3 fatty acid precursor ALA, necessitating the exogenous supplementation of a diet containing n-3 LCPUFAs for optimal health and development [28][29]. In adults, circulating EPA levels increased with DHA supplementation. However, ALA and EPA supplementation did not result in increased DHA levels in circulation [29]. In contrast, Metherel et al. showed that supplemental EPA was converted to DHA [30]. Increased levels of DHA to EPA could be either due to retro-conversion [31][32][33] or reduced EPA metabolism [34], suggesting that circulating levels of EPA and DHA are differently regulated.
A large, prospective, randomized clinical trial of 11,324 patients with a recent myocardial infarction showed a reduced relative risk of death in response to LCPUFAs [35]. A lower rate of myocardial infarction was observed in people consuming an Inuit diet, a diet characterized by the high consumption of marine mammals and fish [36]. The Inuit diet is rich in n-3 LCPUFAs, particularly EPA and DHA compared to those consuming a Western diet, which is abundant in n-6 LCPUFAs, mainly derived from vegetable oils rich in LA [11][37]. Furthermore, a significant difference was observed for cardiovascular mortality in two cohorts with n-3 versus n-6 LCPUFA enriched diets [38][39]. Clinical studies have shown a reduction in mortality of patients with cardiovascular diseases in response to n-3 LCPUFAs [40][41]. Meta-analyses in adults have shown that the Mediterranean diet, enriched in n-3 LCPUFAs [42] and oleic acid [43], has a protective effect against cardiovascular diseases [43][44][45]. These studies declared the importance of bioactive compounds related to the diet. The beneficial effect of n-3 LCPUFAs (EPA and DHA) against cardiovascular diseases could be due to anti-arrhythmic, anti-inflammatory, anti-thrombotic, and hypolipidemic effects and improvement of vascular function [46][47]. However, a recent clinical trial failed to show a protective effect against arrhythmia in patients with myocardial infarction [48]. Supplementation of n-3 LCPUFAs showed no effect on atrial fibrillation [49], cardiac death, myocardial infarction, or stroke [50].
In the Mediterranean diet, one of the main dietary source of n-3 LCPUFAs is fish, particularly oily fish species and shellfish [51]. Neonates, specifically preterm infants, are at risk of a LCPUFA deficit, which has been associated with a subsequent increased risk of various neonatal diseases [52]. Infants with heart disease may require longer parenteral nutrition during their hospitalization that fails to maintain the critical blood levels of LCPUFAs. In neonates, adequate nutritional intake of LCPUFAs is important for visual development, growth, complex brain function, and immune function [53]. Parenteral administration of n-3 LCPUFAs containing lipid emulsion to infants previous to cardiac surgery decreased pro-inflammatory response after surgery [54]. These data suggest the beneficial impact of LCPUFAs in infants with cardiac diseases [52].
2.2. LCPUFAs Derived Mediators and Their Implication in Cardiovascular Physiology
LCPUFAs act as substrates for oxylipin production. LCPUFA metabolism to oxylipins occurs through three different pathways that are mediated by cyclooxygenase (COX), lipoxygenase (LOX), and cytochrome P450 enzymes (CYP) as a result of transcellular biosynthesis and coordination between distinct cell types [55][56][57]. Additionally, oxylipins can be produced by free radical catalyzed non-enzymatic lipid peroxidation [58][59]. High n-6 LCPUFAs and n-3 LCPUFAs intake is associated with higher levels of n-6 and n-3 LCPUFA-derived oxylipins, respectively [55]. In recent years, an active role for to maintain the tissues homeostasis has been attributed to the local generation of specialized pro-resolving mediators (SPMs). SPMs exert their effect by interacting with G-protein coupled receptor (GPCR), such as ALX/FPR2, GPR32, ChemR23, BLT1, and GPR18 [60][61], and modulate diverse biological responses [62] by regulating mitogen-activated protein kinase signaling, NF-κB pathways, AP-1 activation, and oxidative stress-related metabolism [63]. SPMs suppress the expression of adhesion molecules on leukocytes, endothelial cells, neutrophil chemotaxis, and IL-8 production. Frequently, SPMs are rapidly inactivated locally by eicosanoid oxidoreductases and prostaglandin dehydrogenase [64].
Omega-6 mediators. The most well-known oxylipins, the eicosanoids, are formed from ARA. ARA produces series-2 oxylipins via the COX pathway, resulting in the formation of prostaglandin G2 and subsequently to prostaglandin H2, which later gets converted to other prostaglandins and thromboxanes by specific prostaglandin and thromboxane synthases enzymes [65] (Figure 1). Prostaglandins make a huge oxylipins family with multiple biological functions such as inhibition of human platelet aggregation [66][67][68], neutrophil degranulation [69], innate immune response [70], activation of plasminogen activator inhibitor type-1 [71], and reducing pulmonary vascular resistance [72]. Prostaglandins can either have pro-inflammatory and vasoconstrictor effects [73] or anti-inflammatory and vasodilator effects [67][68], depending on to which receptor it binds [74]. Thromboxane A2 is an endothelium-derived oxylipin that potentially induces vasoconstriction and platelet aggregation [67] and thromboxane B2 is positively associated with high central blood pressure and multiple cardiovascular events [75][76].
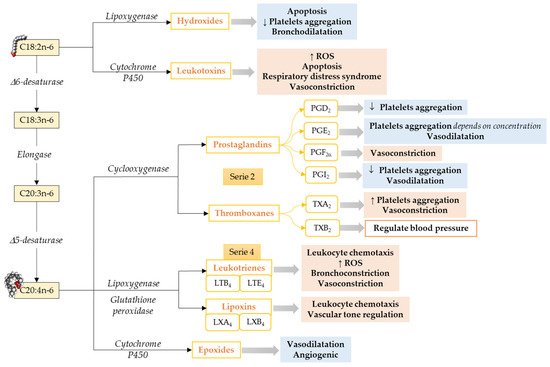
Figure 1. Pathways to synthesize n-6 LCPUFA-derived lipid mediators and their functions focusing on the cardiovascular system and vascular endothelial function. C18:2n-6, Linoleic acid; C18:3n-6, γ-Linolenic acid; C20:3n-6; Dihomo-γ-Linolenic acid; C20:4n-6, Arachidonic acid; ROS, Reactive Oxidative Species. ↓ = Decreases; ↑ = Increases. Modified from [55].
In addition to the COX pathway, oxylipins are also formed by a second pathway involving LOXs that catalyze the formation of hydroxy fatty acid (5-, 12-, and 15-HETE are the most commonly described [77]) and their secondary metabolites, such as leukotrienes, lipoxins, resolvins, protectins, maresins, hepoxilins, and eoxins, via glutathione peroxidase [77]. ARA, via the LOX pathway, produces HpETEs, which can be further converted to leukotrienes (series-4), which are associated with atherosclerosis, endothelial dysfunction, and cytokine release [78]. Moreover, leukotrienes can also be converted to lipoxins [57], which play a role in the resolution of inflammation [79].
The third pathway that leads to oxylipins from LCPUFA metabolism involves a diverse array of membrane-bound cytochrome P450 (CYP) enzymes. ARA metabolism through CYP pathway activity results in the formation of HETE by CYP omega-hydroxylase activity. Cytochrome P450 can also act on ARA to synthesize epoxides, which could have vasodilator and vascular relaxing effects [80].
Limited information is available about LA-derived oxylipins. LA produces oxylipins through the LOX pathway (i.e., 13-HODE), COX pathways (i.e., 9-HODE), and epoxygenase activity of CYP (i.e., EpOME), as well as non-enzymatical pathways (i.e., 9-HODE; Figure 1), and the relative importance of these pathways needs to be explored. LA-derived oxylipins have been shown to attenuate platelet adhesion to endothelial cells [81], induce oxidative stress and a pro-inflammatory response in vascular endothelial cells [82][83], activate plasminogen activator inhibitor type-1 [71], and prevent platelets from adhering to human vascular endothelial cells [84][85]. However, the direct effect of LA-derived oxylipins on the heart has not been studied.
Omega-3 mediators. ALA, a precursor for n-3 LCPUFAs, can be metabolized to oxylipins by LOX [86], COX [55], or CYP cyclooxygenase activity [87] (Figure 2). ALA, metabolized via Cytochrome P450, results in epoxy fatty acids [87], and the levels of these epoxy fatty acids were decreased in adults with hyperlipidemia [88].
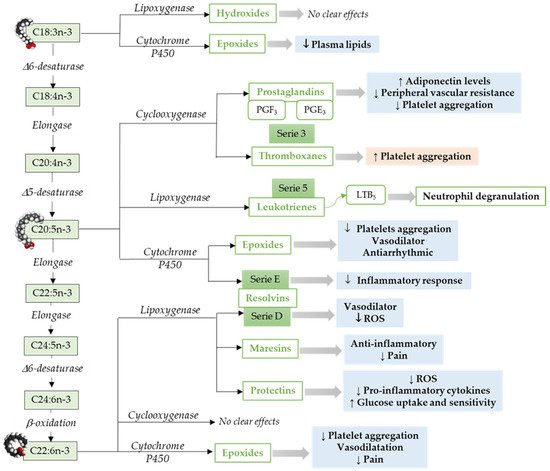
Figure 2. Pathways to synthesize n-3 LCPUFA-derived lipid mediators and its function focusing on the cardiovascular system and vascular endothelial function. C18:3n-3, α-Linolenic acid; C18:4n-3, Stearidonic acid; C20:4n-3, Eicosatetraenoic acid; C20:5n-3, Eicosapensaenoic acid; C22:5n-3, Docosapentaenoic acid; C24:5n-3, Tetracosapentaenoic acid; C24:6n-3, Tetracosahexaenoic acid; C22:5n-3, Docosahexaenoic acid; ROS, Reactive Oxidative Species. ↓ = Decreases; ↑ = Increases. Modified from [55].
Similar to ARA, EPA metabolism through COX pathways generates oxylipins that includes series-3 prostaglandins (i.e., PGE3) and thromboxanes (i.e., thromboxane A3) [55]. However, relative to ARA, EPA is a weak substrate for COX enzyme [89]. Furthermore, EPA via LOX pathway results in leukotrienes (series-5) [90]. An in vitro study has shown that leukotrienes derived from both n-6 and n-3 LCPUFAs induce neutrophil lysosomal degranulation. However, n-3 LCPUFA-derived leukotriene (leukotriene B5) is less effective compared to n-6 LCPUFA-derived leukotriene (leukotriene B4) [91]. In addition, leukotriene B5 could be less inflammatory than leukotrienes 4-series [92]. The epoxides-fatty acids derived from EPA via CYP pathways [93], inhibits platelet aggregation, affect vasodilation, and have an antiarrhythmic effect in neonatal cardiomyocytes [94][95]. E-series resolvins synthesized by cytochrome P450 from EPA [64][96] have been shown to reduce neutrophil migration and inflammatory responses [97].
In addition to EPA, DHA is the other predominant omega-3 LCPUFA. DHA metabolism through LOX pathway leads to the production of maresins, resolvins (D-series), and protectins [55]. These endogenous lipid mediators including lipoxins, among other SPMs [95]. DHA-derived 4-hydroxy-docosahexaenoic acid (14-OH-22:6) potentially inhibits platelet aggregation [98]. Resolvins play an important role in inflammation, vascular biology, and platelet aggregation [99][100]. DHA-derived oxylipins can also be synthesized by COX [101][102] and cytochrome P450 epoxygenase. DHA derived epoxy-fatty acids [93] are known to decrease platelet aggregation and thromboxane A2 synthesis [95][103].
In general, oxylipins generated from n-3 LCPUFAs have lesser effectiveness compared to those derived from n-6 LCPUFAs. The effect of n-3 LCPUFA-derived oxylipins could be antagonized by oxylipins derived from n-6 LCPUFAs [104]. The n-3 LCPUFAs often compete with n-6 LCPUFAs for the same receptor and enzyme. The degree of membrane LCPUFAs incorporation might play an important role in determining the biological effect. Moreover, the enzyme activity could be regulated by increasing or decreasing the initial substrate within a given pathway.
3. The Role of SPMs in Cardiovascular Inflammation Resolution
The resolution of inflammation is tightly regulated by endogenously produced lipid mediators such as SPMs. These molecules can be lipoxins, resolvins, protectins, and maresins [105]. SPMs resolve the inflammation by alleviating the pro-inflammatory response, reducing neutrophil infiltration, and clearing the apoptotic cells through macrophages, and thus, enhancing the host defense. In addition, SPMs restrict the T cells actions, which is the main cellular responses involved in chronic inflammation [106][107]. SPMs are an important modulator of oxidative stress. There is evidence showing lipoxins inhibit leukocyte-dependent generation of reactive species [63]. LXA4 treated cardiomyocytes activate MAP-kinase and Nrf2 pathways [108], whose antioxidant properties are essential for cardiac protection [109]. In animal models, RvD1 reduces reactive oxygen species-mediated IL-1β secretion in macrophages [110] and protects against oxidative stress inflammation inhibiting neutrophil infiltration [111]. RvD1 and maresin 1 have been shown to regulate Nrf2-dependent expression of glutathione peroxidase and superoxide dismutase [112][113]. Kang et al. have reviewed a details summary about the role of EPA- and DHA-derived SPMs and Nrf2-antioxidative responses in cardiac fibrosis [109].
SPMs cause a shift in macrophages from M1 (pro-inflammatory) to M2 (anti-inflammatory) [114][115]. In addition to regulating the innate immune response, SPMs are also important in the adaptive immune response by reducing the NK cells cytotoxicity [116], decreasing memory B cell and antibody production [117]. D-series resolvins and maresin 1 dampen the cytokines production by activated CD8+ T cells, TH1, and TH17 cells and promotes the differentiation of CD4+ T cells into Treg cells, while inhibiting the generation of TH1 and TH17 from naïve CD4+ T cells [106][118]. Patients with chronic heart failure (CHF) have significantly reduced the plasma levels of RvD1 and pretreatment of mononuclear cells of patients with CHF with RvD1 or RvD2, which did not affect cytokine release from CD8+ and CD4+ T cells. This impaired T cells response was associated with reduced GPR32 expression [107]. The interaction between SPMs, their receptors, and their effect depend on the level of SPMs, cell type, and surrounding environment. Uncontrolled and unresolved inflammation can result in cardiovascular diseases, suggesting the critical role of SPMs. An extensive review regarding the lipid signaling pathways in cells and these effects on adult cardiovascular physiology and regeneration was covered by Wasserman et al. [119].
The etiology of cardiovascular disease involves a chronic inflammatory process driven by the formation of lipid-rich lesions in the vascular wall leading to myocardial infarction and stroke. Evidence about the role of SPMs in cardiovascular diseases came from human and animal studies. Serhan et al. have shown increased lipoxin levels in humans after angioplasty [120]. Overexpression of 15-LOX in macrophages resulted in a significant reduction of atherosclerosis in rabbits [121][122], while there was delayed atherogenesis in mice [123]. Administration of RvE1 attenuated atherogenesis in rabbits fed with a high fat and cholesterol diet, possibly by reducing the levels of C-reactive protein (CRP) [124].
The formation of a vulnerable plaque region (a subset of atherosclerotic plaques), with increased inflammation, oxidative stress, and necrotic areas as a result of increased cell death, might lead to acute atherothrombotic clinical events, such as myocardial infarction and stroke, probably due to a defective inflammation-resolution process [125][126][127]. Advance plaque regions in Ldlr−/− or ApoE−/− mice fed a high-fat and high-cholesterol diet displayed an imbalanced ratio of SPMs and pro-inflammatory lipid mediator ratio compared to early plaque regions [128][129]. Furthermore, SPM administration, including RvD2 and Maresin 1, or aspirin-triggered lipoxin A4, caused delayed atherosclerosis and resulted in a more stable-like plaque phenotype [129][130], probably due to decreased necrosis, oxidative stress, and increased fibrous cap thickness and helping in tissue repair processes.
Decreased blood flow can lead to tissue injury, as a result of the inflammatory response, due to leukocyte infiltration and ROS production [131]. This suggests that controlled leukocyte infiltration, as well as their removal from the site of injury, is important to protect the heart and myocardium during ischemia. Indeed, in mice, exogenous administration of RvD1 has been shown to improve cardiac function by reducing leukocyte infiltration and fibrosis [132]. Moreover, in a rat model of myocardial ischemia/reperfusion injury, RvE1 protects the rat heart by decreasing infarct size [133]. In addition to the heart, the protective effects of SPMs have been reported in other tissues, including the kidney and lung [134][135], possibly by reducing leukocyte recruitment.
Atherothrombosis is one of the clinical manifestations which leads to myocardial infarction and stroke [136]. Platelets and neutrophils aggregation are important for plaque inflammation [137]. Increased levels of circulating leukocyte-platelets aggregates in cardiovascular diseases suggest their contribution to pathogenesis. Moreover, elevated levels of platelet–monocyte aggregates have been used as an early marker of acute myocardial infarction [138]. It is possible that platelets and neutrophils/macrophage aggregates cause plaque formation and cardiovascular disease and are unable to produce enough SPMs. The levels of lipid mediators, specifically thromboxane and prostacyclin, are critical for platelet activation and thrombosis. In a randomized human-clinical trial, healthy subjects receiving aspirin at a recommended dose for patients with cardiovascular diseases (low dose = 81 mg) resulted in aspirin-induced 15-epi-LXA4, and the levels were negatively correlated with plasma thromboxane B2 levels, reducing the platelets activation [139]. Moreover, EPA-derived RvE1 has been shown to have an antiplatelet aggregation activity [140]. In response to acute inflammation, the self-limiting events such as platelets and neutrophils aggregation are critical and involve the biosynthesis of SPM to timely resolve the inflammation by reducing platelets and neutrophils aggregation, decrease cytokines production, and increase apoptosis [141][142].
SPMs, such as marsein 1, induce a pro-resolving platelet phenotype by increasing platelet aggregation and decreasing the levels of pro-inflammatory and pro-thrombotic mediators. All these data suggest the key role of SPM in promoting resolution inflammation as well as thrombosis during inflammation, and thus, could have potential therapeutic implications in cardiovascular diseases.
SPMs in Infant Cardiovascular Health and Disease
LCPUFAs and oxylipins can modulate the balance for infant cardiac health and disease, by regulating the inflammation pathways. Gestational age and birth weight are important risk factors for the development of cardiovascular diseases [3][143][144], possibly by reducing endothelial function [145][146]. Prostaglandin E2 treatment is widely used in aortic coarctation [147], which is the third most common congenital cardiac lesion in preterm infants [148], by widening of the constricted ductal tissue within the aorta. Conversely, prostaglandin E1 in near-term infants resulted in a worsening of the aortic constriction [149]. Differences in pharmacokinetics and pharmacodynamics efficacy of prostaglandin therapy depend on the clinical course, the effects of surfactant-deficient lung disease, concurrent infections, myocardial insufficiency, and hemodynamic instability.
The ductus arteriosus is the fetal artery that connects the pulmonary artery and the aorta. The closure of the ductus arteriosus mostly occurs within three days of life in healthy term newborns as a result of increased oxygen level and reduced prostaglandin E2 [150]. However, in preterm infants, the ductus fails to close, resulting in patent ductus arteriosus (PDA). PDA is a heart problem that is common in preterm infants and causes morbidities such as bronchopulmonary dysplasia, intraventricular hemorrhage, and necrotizing enterocolitis and mortality [151][152]. The current strategy to treat ductus arteriosus involves the use of prostaglandins [151].
Inhibition of ARA-derived epoxides hydrolases showed beneficial cardiovascular effects, including vasodilation, anti-inflammation, anti-hypertrophy, and myocardial protection [153][154]. However, other data showed that epoxide hydrolase inhibition did not prevent cardiac remodeling or dysfunction [155], suggesting that targeting particular oxylipins seems to be insufficient for preventing cardiovascular events.
The E-series and D-series resolvins derived from n-3 fatty acids are important mediators in the resolution of inflammation [156], but their ability to modulate contraction of vascular smooth muscle is not known. In cardiovascular diseases, resolvins E1, D1, and D2 prevented constriction in human arteries induced by thromboxane [157] by resolving inflammation and preventing inappropriate vascular contractility.
Altered levels of resolvins are associated with cardiovascular disease onset, propagation, and systemic inflammation. Plasma levels of D-series resolvins were negatively correlated with decreased platelets and leukocyte activation [158]. Resolvin D1 administration in mice resulted in improved ventricular function following myocardial infarction by activating inflammatory response [132]. Most of the human studies regarding the effect of SPMs in cardiovascular disease have been done either in adult population or in children over 3 years. How these oxylipins act in neonatal cardiovascular remodeling and how nutritional intervention modulates this interaction need to be explored.
References
- Bansilal, S.; Castellano, J.M.; Fuster, V. Global burden of CVD: Focus on secondary prevention of cardiovascular disease. Int. J. Cardiol. 2015, 201, S1–S7.
- Swirski, F.K.; Nahrendorf, M. Leukocyte Behavior in Atherosclerosis, Myocardial Infarction, and Heart Failure. Science 2013, 339, 161–166.
- Visentin, S.; Grumolato, F.; Nardelli, G.B.; Di Camillo, B.; Grisan, E.; Cosmi, E. Early origins of adult disease: Low birth weight and vascular remodeling. Atherosclerosis 2014, 237, 391–399.
- Kris-Etherton, P.M.; Etherton, T.D.; Carlson, J.; Gardner, C. Recent discoveries in inclusive food-based approaches and dietary patterns for reduction in risk for cardiovascular disease. Curr. Opin. Lipidol. 2002, 13, 397–407.
- Hasler, C.M.; Kundrat, S.; Wool, D. Functional foods and cardiovascular disease. Curr. Atheroscler. Rep. 2000, 2, 467–475.
- Nathan, C.; Ding, A. Nonresolving Inflammation. Cell 2010, 140, 871–882.
- Chiurchiù, V.; Leuti, A.; Maccarrone, M. Bioactive Lipids and Chronic Inflammation: Managing the Fire Within. Front. Immunol. 2018, 9, 38.
- Shapiro, M.D.; Fazio, S. From Lipids to Inflammation. Circ. Res. 2016, 118, 732–749.
- Khan, S.U.; Khan, M.S.; Riaz, H.; Valavoor, S.; Zhao, D.; Vaughan, L.; Okunrintemi, V.; bin Riaz, I.; Kaluski, E.; Murad, M.H.; et al. Effects of Nutritional Supplements and Dietary Interventions on Cardiovascular Outcomes. Ann. Intern. Med. 2019, 171, 190–198.
- Fredman, G.; Spite, M. Specialized pro-resolving mediators in cardiovascular diseases. Mol. Asp. Med. 2017, 58, 65–71.
- Schunck, W.-H.; Konkel, A.; Fischer, R.; Weylandt, K.-H. Therapeutic potential of omega-3 fatty acid-derived epoxyeicosanoids in cardiovascular and inflammatory diseases. Pharmacol. Ther. 2018, 183, 177–204.
- De Carvalho, A.E.T.S.; Bassaneze, V.; Forni, M.F.; Keusseyan, A.A.; Kowaltowski, A.J.; Krieger, J.E. Early Postnatal Cardiomyocyte Proliferation Requires High Oxidative Energy Metabolism. Sci. Rep. 2017, 7, 15434.
- Porrello, E.R.; Mahmoud, A.I.; Simpson, E.; Hill, J.A.; Richardson, J.A.; Olson, E.N.; Sadek, H. Transient Regenerative Potential of the Neonatal Mouse Heart. Science 2011, 331, 1078–1080.
- Zogbi, C.; De Carvalho, A.E.T.S.; Nakamuta, J.S.; Caceres, V.D.M.; Prando, S.; Giorgi, M.C.P.; Rochitte, C.E.; Meneghetti, J.C.; Krieger, J.E. Early postnatal rat ventricle resection leads to long-term preserved cardiac function despite tissue hypoperfusion. Physiol. Rep. 2014, 2, e12115.
- Soonpaa, M.H.; Kim, K.K.; Pajak, L.; Franklin, M.; Field, L.J. Cardiomyocyte DNA synthesis and binucleation during murine development. Am. J. Physiol. Circ. Physiol. 1996, 271, H2183–H2189.
- Cao, T.; Liccardo, D.; Lacanna, R.; Zhang, X.; Lu, R.; Finck, B.N.; Leigh, T.; Chen, X.; Drosatos, K.; Tian, Y. Fatty Acid Oxidation Promotes Cardiomyocyte Proliferation Rate but Does Not Change Cardiomyocyte Number in Infant Mice. Front. Cell Dev. Biol. 2019, 7.
- Lopaschuk, G.D.; Jaswal, J.S. Energy Metabolic Phenotype of the Cardiomyocyte During Development, Differentiation, and Postnatal Maturation. J. Cardiovasc. Pharmacol. 2010, 56, 130–140.
- Zebrowski, D.C.; Engel, F.B. The Cardiomyocyte Cell Cycle in Hypertrophy, Tissue Homeostasis, and Regeneration. Rev. Physiol. Biochem. Pharmacol. 2013, 165, 67–96.
- Makinde, A.O.; Kantor, P.F.; Lopaschuk, G.D. Maturation of fatty acid and carbohydrate metabolism in the newborn heart. Mol. Cell Biochem. 1998, 188, 49–56.
- Vishwanath, V.A. Fatty Acid Beta-Oxidation Disorders: A Brief Review. Ann. Neurosci. 2016, 23, 51–55.
- Miklas, J.W.; Clark, E.; Levy, S.; Detraux, D.; Leonard, A.; Beussman, K.; Showalter, M.R.; Smith, A.T.; Hofsteen, P.; Yang, X.; et al. TFPa/HADHA is required for fatty acid beta-oxidation and cardiolipin re-modeling in human cardiomyocytes. Nat. Commun. 2019, 10, 1–21.
- Puente, B.N.; Kimura, W.; Muralidhar, S.A.; Moon, J.; Amatruda, J.F.; Phelps, K.L.; Grinsfelder, D.; Rothermel, B.A.; Chen, R.; Garcia, J.A.; et al. The Oxygen-Rich Postnatal Environment Induces Cardiomyocyte Cell-Cycle Arrest through DNA Damage Response. Cell 2014, 157, 565–579.
- Delbridge, L.M.D.; Mellor, K.M.; Taylor, D.J.R.; Gottlieb, R.A. Myocardial autophagic energy stress responses--macroautophagy, mitophagy, and glycophagy. Am. J. Physiol. Circ. Physiol. 2015, 308, H1194–H1204.
- Zhang, M.; Mileykovskaya, E.; Dowhan, W. Cardiolipin Is Essential for Organization of Complexes III and IV into a Supercomplex in Intact Yeast Mitochondria. J. Biol. Chem. 2005, 280, 29403–29408.
- Chatfield, K.C.; Sparagna, G.C.; Sucharov, C.C.; Miyamoto, S.D.; Grudis, J.E.; Sobus, R.D.; Hijmans, J.; Stauffer, B.L. Dysregulation of cardiolipin biosynthesis in pediatric heart failure. J. Mol. Cell. Cardiol. 2014, 74, 251–259.
- Sparagna, G.C.; Chicco, A.J.; Murphy, R.C.; Bristow, M.R.; Johnson, C.A.; Rees, M.L.; Maxey, M.L.; McCune, S.A.; Moore, R.L. Loss of cardiac tetralinoleoyl cardiolipin in human and experimental heart failure. J. Lipid Res. 2007, 48, 1559–1570.
- Mulligan, C.M.; Sparagna, G.C.; Le, C.H.; De Mooy, A.B.; Routh, M.A.; Holmes, M.G.; Hickson-Bick, D.L.; Zarini, S.; Murphy, R.C.; Xu, F.Y.; et al. Dietary linoleate preserves cardiolipin and attenuates mitochondrial dysfunction in the failing rat heart. Cardiovasc. Res. 2012, 94, 460–468.
- Zárate, R.; El Jaber-Vazdekis, N.; Tejera, N.; Pérez, J.A.P.; Rodríguez, C. Significance of long chain polyunsaturated fatty acids in human health. Clin. Transl. Med. 2017, 6, 25.
- Brenna, J.T.; Salem, N.; Sinclair, A.J.; Cunnane, S.C. α-Linolenic acid supplementation and conversion to n-3 long-chain polyunsaturated fatty acids in humans. Prostaglandins Leukot. Essent. Fat. Acids 2009, 80, 85–91.
- Metherel, A.H.; Irfan, M.; Klingel, S.L.; Mutch, D.M.; Bazinet, R.P. Compound-specific isotope analysis reveals no retroconversion of DHA to EPA but substantial conversion of EPA to DHA following supplementation: A randomized control trial. Am. J. Clin. Nutr. 2019, 110, 823–831.
- Conquer, J.A.; Holub, B.J. Supplementation with an Algae Source of Docosahexaenoic Acid Increases (n-3) Fatty Acid Status and Alters Selected Risk Factors for Heart Disease in Vegetarian Subjects. J. Nutr. 1996, 126, 3032–3039.
- Brossard, N.; Croset, M.; Pachiaudi, C.; Riou, J.P.; Tayot, J.L.; Lagarde, M. Retroconversion and metabolism of [13C]22:6n−3 in humans and rats after intake of a single dose of [13C]22:6n−3-triacylglycerols. Am. J. Clin. Nutr. 1996, 64, 577–586.
- Conquer, J.A.; Holub, B.J. Dietary docosahexaenoic acid as a source of eicosapentaenoic acid in vegetarians and omnivores. Lipids 1997, 32, 341–345.
- Plourde, M.; Chouinard-Watkins, R.; Vandal, M.; Zhang, Y.; Lawrence, P.; Brenna, J.T.; Cunnane, S.C. Plasma incorporation, apparent retroconversion and β-oxidation of 13C-docosahexaenoic acid in the elderly. Nutr. Metab. 2011, 8, 5.
- Gruppo Italiano per lo Studio della Sopravvivenza nell’Infarto miocardico. Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: Results of the GISSI-Prevenzione trial. Lancet 1999, 354, 447–455.
- Hu, X.F.; Kenny, T.-A.; Chan, H.M. Inuit Country Food Diet Pattern Is Associated with Lower Risk of Coronary Heart Disease. J. Acad. Nutr. Diet. 2018, 118, 1237–1248.e1.
- Dyerberg, J.; Bang, H.; Stoffersen, E.; Moncada, S.; Vane, J. Eicosapentaenoic acid and prevention of thrombosis and atherosclerosis? Lancet 1978, 312, 117–119.
- Lands, W.E. Dietary Fat and Health: The Evidence and the Politics of Prevention: Careful Use of Dietary Fats Can Improve Life and Prevent Disease. Ann. N. Y. Acad. Sci. 2005, 1055, 179–192.
- Von Schacky, C.; Harris, W.S. Cardiovascular benefits of omega-3 fatty acids. Cardiovasc. Res. 2007, 73, 310–315.
- Marchioli, R.; Barzi, F.; Bomba, E.; Chieffo, C.; Di Gregorio, D.; Di Mascio, R.; Franzosi, M.G.; Geraci, E.; Levantesi, G.; Maggioni, A.P.; et al. Early Protection Against Sudden Death by n-3 Polyunsaturated Fatty Acids After Myocardial Infarction. Circulation 2002, 105, 1897–1903.
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067.
- Román, G.C.; Jackson, R.E.; Gadhia, R.; Román, A.N.; Reis, J. Mediterranean diet: The role of long-chain ω-3 fatty acids in fish; polyphenols in fruits, vegetables, cereals, coffee, tea, cacao and wine; probiotics and vitamins in prevention of stroke, age-related cognitive decline, and Alzheimer disease. Rev. Neurol. 2019, 175, 724–741.
- Sales-Campos, H.; Souza, P.R.; Peghini, B.C.; da Silva, J.S.; Cardoso, C.R. An overview of the modulatory effects of oleic acid in health and disease. Mini Rev. Med. Chem. 2013, 13, 201–210.
- Widmer, R.J.; Flammer, A.J.; Lerman, L.O.; Lerman, A. The Mediterranean Diet, its Components, and Cardiovascular Disease. Am. J. Med. 2015, 128, 229–238.
- Liyanage, T.; Ninomiya, T.; Wang, A.; Neal, B.; Jun, M.; Wong, M.G.; Jardine, M.; Hillis, G.S.; Perkovic, V. Effects of the Mediterranean Diet on Cardiovascular Outcomes—A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0159252.
- Lavie, C.J.; Milani, R.V.; Mehra, M.R.; Ventura, H.O. Omega-3 Polyunsaturated Fatty Acids and Cardiovascular Diseases. J. Am. Coll. Cardiol. 2009, 54, 585–594.
- Mozaffarian, D. Fish and n−3 fatty acids for the prevention of fatal coronary heart disease and sudden cardiac death. Am. J. Clin. Nutr. 2008, 87, 1991S–1996S.
- Rauch, B.; Schiele, R.; Schneider, S.; Diller, F.; Victor, N.; Gohlke, H.; Gottwik, M.; Steinbeck, G.; Del Castillo, U.; Sack, R.; et al. OMEGA, a Randomized, Placebo-Controlled Trial to Test the Effect of Highly Purified Omega-3 Fatty Acids on Top of Modern Guideline-Adjusted Therapy After Myocardial Infarction. Circulation 2010, 122, 2152–2159.
- Martino, A.; Pezzi, L.; Magnano, R.; Salustri, E.; Penco, M.; Calo’, L. Omega 3 and atrial fibrillation: Where are we? World J. Cardiol. 2016, 8, 114–119.
- Rizos, E.C.; Ntzani, E.E.; Bika, E.; Kostapanos, M.S.; Elisaf, M.S. Association Between Omega-3 Fatty Acid Supplementation and Risk of Major Cardiovascular Disease Events. JAMA 2012, 308, 1024–1033.
- Hepburn, F.N.; Exler, J.; Weihrauch, J.L. Provisional tables on the content of omega-3 fatty acids and other fat components of selected foods. J. Am. Diet. Assoc. 1986, 86, 788–793.
- Lewis, K.D.; Conway, J.; Cunningham, C.; Larsen, B.M.K. Optimizing Nutrition in Pediatric Heart Failure: The Crisis Is Over and Now It’s Time to Feed. Nutr. Clin. Pr. 2017, 33, 397–403.
- Valentová, M.; Von Haehling, S.; Doehner, W.; Murín, J.; Anker, S.D.; Sandek, A. Liver dysfunction and its nutritional implications in heart failure. Nutrients 2013, 29, 370–378.
- Larsen, B.M.K.; Field, C.J.; Leong, A.Y.; Goonewardene, L.A.; Van Aerde, J.E.; Joffe, A.R.; Clandinin, M.T. Pretreatment with an Intravenous Lipid Emulsion Increases Plasma Eicosapentanoic Acid and Downregulates Leukotriene B4, Procalcitonin, and Lymphocyte Concentrations after Open Heart Surgery in Infants. J. Parenter. Enter. Nutr. 2013, 39, 171–179.
- Gabbs, M.; Leng, S.; Devassy, J.G.; Monirujjaman, M.; Aukema, H.M. Advances in Our Understanding of Oxylipins Derived from Dietary PUFAs. Adv. Nutr. 2015, 6, 513–540.
- Sala, A.; Folco, G.; Murphy, R.C. Transcellular biosynthesis of eicosanoids. Pharmacol. Rep. 2010, 62, 503–510.
- Bannenberg, G.; Serhan, C.N. Specialized pro-resolving lipid mediators in the inflammatory response: An update. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2010, 1801, 1260–1273.
- Niki, E.; Yoshida, Y. Biomarkers for oxidative stress: Measurement, validation, and application. J. Med. Investig. 2005, 52, 228–230.
- Barbosa, M.; Valentão, P.; Andrade, P.B. Biologically Active Oxylipins from Enzymatic and Nonenzymatic Routes in Macroalgae. Mar. Drugs 2016, 14, 23.
- Serhan, C.N. Pro-resolving lipid mediators are leads for resolution physiology. Nature 2014, 510, 92–101.
- Chiang, N.; Serhan, C.N.; Dahlén, S.-E.; Drazen, J.M.; Hay, D.W.P.; Rovati, G.E.; Shimizu, T.; Yokomizo, T.; Brink, C. The Lipoxin Receptor ALX: Potent Ligand-Specific and Stereoselective Actions in Vivo. Pharmacol. Rev. 2006, 58, 463–487.
- McMahon, B.; Godson, C. Lipoxins: Endogenous regulators of inflammation. Am. J. Physiol. Physiol. 2004, 286, F189–F201.
- Leuti, A.; Maccarrone, M.; Chiurchiù, V. Proresolving Lipid Mediators: Endogenous Modulators of Oxidative Stress. Oxid. Med. Cell. Longev. 2019, 2019, 1–12.
- Conte, M.S.; Desai, T.A.; Wu, B.; Schaller, M.; Werlin, E. Pro-resolving lipid mediators in vascular disease. J. Clin. Investig. 2018, 128, 3727–3735.
- Smith, W.L.; Urade, Y.; Jakobsson, P.-J. Enzymes of the Cyclooxygenase Pathways of Prostanoid Biosynthesis. Chem. Rev. 2011, 111, 5821–5865.
- Meydani, S.N.; Shapiro, A.C.; Meydani, M.; Blumberg, J.B. Lung Eicosanoid Synthesis is Affected by Age, Dietary Fat and Vitamin E. J. Nutr. 1992, 122, 1627–1633.
- Cheng, Y.; Austin, S.C.; Rocca, B.; Koller, B.H.; Coffman, T.M.; Grosser, T.; Lawson, J.A.; Fitzgerald, G.A. Role of Prostacyclin in the Cardiovascular Response to Thromboxane A2. Science 2002, 296, 539–541.
- Dusting, G.J.; Moncada, S.; Vane, J.R. Prostacyclin (PGX) is the endogeneous metabolite responsible for relaxation of coronary arteries induced by arachidonic acid. Prostaglandins 1977, 13, 3–15.
- Darius, H.; Michael-Hepp, J.; Thierauch, K.-H.; Fisch, A. Inhibition of human platelets and polymorphonuclear neutrophils by the potent and metabolically stable prostaglandin D2 analog ZK 118.182. Eur. J. Pharmacol. 1994, 258, 207–213.
- Seubert, J.; Yang, B.; Bradbury, J.A.; Graves, J.; DeGraff, L.M.; Gabel, S.; Gooch, R.; Foley, J.; Newman, J.; Mao, L.; et al. Enhanced Postischemic Functional Recovery in CYP2J2 Transgenic Hearts Involves Mitochondrial ATP-Sensitive K + Channels and p42/p44 MAPK Pathway. Circ. Res. 2004, 95, 506–514.
- Marmol, F.; Puig-Parellada, P.; Sanchez, J.; Trullas, R. Influence of aging on thromboxane A2 and prostacyclin levels in rat hippocampal brain slices. Neurobiol. Aging 1999, 20, 695–697.
- Matz, R.L.; De Sotomayor, M.A.; Schott, C.; Stoclet, J.-C.; Andriantsitohaina, R. Vascular bed heterogeneity in age-related endothelial dysfunction with respect to NO and eicosanoids. Br. J. Pharmacol. 2000, 131, 303–311.
- Kuhn, H.; Banthiya, S.; Van Leyen, K. Mammalian lipoxygenases and their biological relevance. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2015, 1851, 308–330.
- A Coleman, R.; Smith, W.L.; Narumiya, S. International Union of Pharmacology classification of prostanoid receptors: Properties, distribution, and structure of the receptors and their subtypes. Pharmacol. Rev. 1994, 46, 205–229.
- Caligiuri, S.P.; Aukema, H.M.; Ravandi, A.; Lavallée, R.; Guzman, R.; Pierce, G.N. Specific plasma oxylipins increase the odds of cardiovascular and cerebrovascular events in patients with peripheral artery disease. Can. J. Physiol. Pharmacol. 2017, 95, 961–968.
- Caligiuri, S.P.; Rodriguez-Leyva, D.; Aukema, H.M.; Ravandi, A.; Weighell, W.; Guzman, R.; Pierce, G.N. Dietary Flaxseed Reduces Central Aortic Blood Pressure Without Cardiac Involvement but Through Changes in Plasma Oxylipins. Hypertension 2016, 68, 1031–1038.
- Sutherland, M.; Shankaranarayanan, P.; Schewe, T.; Nigam, S. Evidence for the presence of phospholipid hydroperoxide glutathione peroxidase in human platelets: Implications for its involvement in the regulatory network of the 12-lipoxygenase pathway of arachidonic acid metabolism. Biochem. J. 2001, 353, 91–100.
- Bäck, M. Leukotriene Signaling in Atherosclerosis and Ischemia. Cardiovasc. Drugs Ther. 2008, 23, 41–48.
- Buckley, C.D.; Gilroy, D.W.; Serhan, C.N. Proresolving Lipid Mediators and Mechanisms in the Resolution of Acute Inflammation. Immunity 2014, 40, 315–327.
- Alsaad, A.M.S.; Zordoky, B.; Tse, M.M.Y.; El-Kadi, A.O.S. Role of cytochrome P450–mediated arachidonic acid metabolites in the pathogenesis of cardiac hypertrophy. Drug Metab. Rev. 2013, 45, 173–195.
- Tloti, M.; Moon, D.; Weston, L.; Kaplan, J. Effect of 13-hydroxyoctadeca-9,11-dienoic acid (13-hode) on thrombin induced platelet adherence to endothelial cells in vitro. Thromb. Res. 1991, 62, 305–317.
- Ferdouse, A.; Leng, S.; Winter, T.; Aukema, H. Dietary n-6 and n-3 PUFA alter the free oxylipin profile differently in male and female rat hearts. Br. J. Nutr. 2019, 122, 252–261.
- Viswanathan, S.; Hammock, B.D.; Newman, J.W.; Meerarani, P.; Toborek, M.; Hennig, B. Involvement of CYP 2C9 in mediating the proinflammatory effects of linoleic acid in vascular endothelial cells. J. Am. Coll. Nutr. 2003, 22, 502–510.
- Theken, K.; Schuck, R.N.; Edin, M.L.; Tran, B.; Ellis, K.; Bass, A.; Lih, F.B.; Tomer, K.B.; Poloyac, S.M.; Wu, M.C.; et al. Evaluation of cytochrome P450-derived eicosanoids in humans with stable atherosclerotic cardiovascular disease. Atherosclerosis 2012, 222, 530–536.
- Buchanan, M.R.; A Haas, T.; Lagarde, M.; Guichardant, M. 13-Hydroxyoctadecadienoic acid is the vessel wall chemorepellant factor, LOX. J. Biol. Chem. 1985, 260, 16056–16059.
- Liu, M.; Chen, P.; Véricel, E.; Lelli, M.; Béguin, L.; Lagarde, M.; Guichardant, M. Characterization and biological effects of di-hydroxylated compounds deriving from the lipoxygenation of ALA. J. Lipid Res. 2013, 54, 2083–2094.
- Laethem, R.M.; Balazy, M.; Koop, D.R. Epoxidation of C18 unsaturated fatty acids by cytochromes P4502C2 and P4502CAA. Drug Metab. Dispos. 1996, 24, 664–668.
- Schuchardt, J.P.; Schmidt, S.; Kressel, G.; Dong, H.; Willenberg, I.; Hammock, B.D.; Hahn, A.; Schebb, N.H. Comparison of free serum oxylipin concentrations in hyper- vs. normolipidemic men. Prostaglandins Leukot. Essent. Fat. Acids 2013, 89, 19–29.
- Wada, M.; DeLong, C.J.; Hong, Y.H.; Rieke, C.J.; Song, I.; Sidhu, R.S.; Yuan, C.; Warnock, M.; Schmaier, A.H.; Yokoyama, C.; et al. Enzymes and Receptors of Prostaglandin Pathways with Arachidonic Acid-derived Versus Eicosapentaenoic Acid-derived Substrates and Products*. J. Biol. Chem. 2007, 282, 22254–22266.
- Hersberger, M. Potential role of the lipoxygenase derived lipid mediators in atherosclerosis: Leukotrienes, lipoxins and resolvins. Clin. Chem. Lab. Med. 2010, 48, 1063–1073.
- Goldman, D.; Pickett, W.; Goetzl, E. Human neutrophil chemotactic and degranulating activities of leukotriene B5 (LTB5) derived from eicosapentaenoic acid. Biochem. Biophys. Res. Commun. 1983, 117, 282–288.
- Mickleborough, T.D.; Lindley, M.R.; Ionescu, A.A.; Fly, A.D. Protective Effect of Fish Oil Supplementation on Exercise-Induced Bronchoconstriction in Asthma. Chest 2006, 129, 39–49.
- Fer, M.; Dréano, Y.; Lucas, D.; Corcos, L.; Salaün, J.-P.; Berthou, F.; Amet, Y. Metabolism of eicosapentaenoic and docosahexaenoic acids by recombinant human cytochromes P450. Arch. Biochem. Biophys. 2008, 471, 116–125.
- Yang, C.; Yang, J.; Xu, X.; Yan, S.; Pan, S.; Pan, X.; Zhang, C.; Leung, G.P. Vasodilatory effect of 14,15-epoxyeicosatrienoic acid on mesenteric arteries in hypertensive and aged rats. Prostaglandins Other Lipid Mediat. 2014, 112, 1–8.
- Caligiuri, S.P.B.; Parikh, M.; Stamenkovic, A.; Pierce, G.N.; Aukema, H.M. Dietary modulation of oxylipins in cardiovascular disease and aging. Am. J. Physiol. Circ. Physiol. 2017, 313, H903–H918.
- Isobe, Y.; Arita, M.; Matsueda, S.; Iwamoto, R.; Fujihara, T.; Nakanishi, H.; Taguchi, R.; Masuda, K.; Sasaki, K.; Urabe, D.; et al. Identification and Structure Determination of Novel Anti-inflammatory Mediator Resolvin E3, 17,18-Dihydroxyeicosapentaenoic Acid*. J. Biol. Chem. 2012, 287, 10525–10534.
- Oh, S.F.; Pillai, P.S.; Recchiuti, A.; Yang, R.; Serhan, C.N. Pro-resolving actions and stereoselective biosynthesis of 18S E-series resolvins in human leukocytes and murine inflammation. J. Clin. Investig. 2011, 121, 569–581.
- Croset, M.; Sala, A.; Folco, G.; Lagarde, M. Inhibition by lipoxygenase products of TXA2-like responses of platelets and vascular smooth muscle. Biochem. Pharmacol. 1988, 37, 1275–1280.
- Sansbury, B.E.; Spite, M. Resolution of Acute Inflammation and the Role of Resolvins in Immunity, Thrombosis, and Vascular Biology. Circ. Res. 2016, 119, 113–130.
- Chen, P.; Vãricel, E.; Lagarde, M.; Guichardant, M. Poxytrins, a class of oxygenated products from polyunsaturated fatty acids, potently inhibit blood platelet aggregation. FASEB J. 2010, 25, 382–388.
- Serhan, C.N.; Hong, S.; Gronert, K.; Colgan, S.P.; Devchand, P.R.; Mirick, G.; Moussignac, R.-L. Resolvins. J. Exp. Med. 2002, 196, 1025–1037.
- Shinohara, M.; Mirakaj, V.; Serhan, C.N. Functional Metabolomics Reveals Novel Active Products in the DHA Metabolome. Front. Immunol. 2012, 3, 81.
- Johnson, E.N.; Brass, L.F.; Funk, C.D. Increased platelet sensitivity to ADP in mice lacking platelet-type 12-lipoxygenase. Proc. Natl. Acad. Sci. USA 1998, 95, 3100–3105.
- Tull, S.P.; Yates, C.M.; Maskrey, B.H.; O’Donnell, V.B.; Madden, J.; Grimble, R.F.; Calder, P.C.; Nash, G.B.; Rainger, G.E. Omega-3 Fatty Acids and Inflammation: Novel Interactions Reveal a New Step in Neutrophil Recruitment. PLoS Biol. 2009, 7, e1000177.
- Serhan, C.N.; Petasis, N.A. Resolvins and Protectins in Inflammation Resolution. Chem. Rev. 2011, 111, 5922–5943.
- Chiurchiù, V.; Leuti, A.; Dalli, J.; Jacobsson, A.; Battistini, L.; Maccarrone, M.; Serhan, C.N. Proresolving lipid mediators resolvin D1, resolvin D2, and maresin 1 are critical in modulating T cell responses. Sci. Transl. Med. 2016, 8, 353ra111.
- Chiurchiù, V.; Leuti, A.; Saracini, S.; Fontana, D.; Finamore, P.; Giua, R.; Padovini, L.; Incalzi, R.A.; Maccarrone, M. Resolution of inflammation is altered in chronic heart failure and entails a dysfunctional responsiveness of T lymphocytes. FASEB J. 2018, 33, 909–916.
- Chen, X.-Q.; Wu, S.-H.; Zhou, Y.; Tang, Y.-R. Lipoxin A4-Induced Heme Oxygenase-1 Protects Cardiomyocytes against Hypoxia/Reoxygenation Injury via p38 MAPK Activation and Nrf2/ARE Complex. PLoS ONE 2013, 8, e67120.
- Kang, G.J.; Kim, E.J.; Lee, C.H. Therapeutic Effects of Specialized Pro-Resolving Lipids Mediators on Cardiac Fibrosis via NRF2 Activation. Antioxidants 2020, 9, 1259.
- Cox, R.; Phillips, O.; Fukumoto, J.; Fukumoto, I.; Parthasarathy, P.T.; Mandry, M.; Cho, Y.; Lockey, R.F.; Kolliputi, N. Resolvins Decrease Oxidative Stress Mediated Macrophage and Epithelial Cell Interaction through Decreased Cytokine Secretion. PLoS ONE 2015, 10, e0136755.
- Spite, M.; Summers, L.; Porter, T.; Srivastava, S.; Bhatnagar, A.; Serhan, C. Resolvin D1 controls inflammation initiated by glutathione-lipid conjugates formed during oxidative stress. Br. J. Pharmacol. 2009, 158, 1062–1073.
- Zhao, Q.; Wu, J.; Lin, Z.; Hua, Q.; Zhang, W.; Ye, L.; Wu, G.; Du, J.; Xia, J.; Chu, M.; et al. Resolvin D1 Alleviates the Lung Ischemia Reperfusion Injury via Complement, Immunoglobulin, TLR4, and Inflammatory Factors in Rats. Inflammation 2016, 39, 1319–1333.
- Sun, Q.; Wu, Y.; Zhao, F.; Wang, J. Maresin 1 Ameliorates Lung Ischemia/Reperfusion Injury by Suppressing Oxidative Stress via Activation of the Nrf-2-Mediated HO-1 Signaling Pathway. Oxid. Med. Cell. Longev. 2017, 2017, 1–12.
- Prieto, P.; Cuenca, J.; Traves, P.G.; Fernández-Velasco, M.; Martín-Sanz, P.; Boscá, L. Lipoxin A4 impairment of apoptotic signaling in macrophages: Implication of the PI3K/Akt and the ERK/Nrf-2 defense pathways. Cell Death Differ. 2010, 17, 1179–1188.
- Börgeson, E.; Johnson, A.M.; Lee, Y.S.; Till, A.; Syed, G.; Ali-Shah, S.T.; Guiry, P.J.; Dalli, J.; Colas, R.A.; Serhan, C.N.; et al. Lipoxin A4 Attenuates Obesity-Induced Adipose Inflammation and Associated Liver and Kidney Disease. Cell Metab. 2015, 22, 125–137.
- Ramstedt, U.; Serhan, C.N.; Nicolaou, K.C.; E Webber, S.; Wigzell, H.; Samuelsson, B. Lipoxin A-induced inhibition of human natural killer cell cytotoxicity: Studies on stereospecificity of inhibition and mode of action. J. Immunol. 1987, 138, 266–270.
- Ramon, S.; Bancos, S.; Serhan, C.N.; Phipps, R.P. LipoxinA4modulates adaptive immunity by decreasing memoryB-cell responses via anALX/FPR2-dependent mechanism. Eur. J. Immunol. 2014, 44, 357–369.
- Pauter, A.M.; Olsson, P.; Asadi, A.; Herslöf, B.; Csikasz, R.I.; Zadravec, D.; Jacobsson, A. Elovl2 ablation demonstrates that systemic DHA is endogenously produced and is essential for lipid homeostasis in mice. J. Lipid Res. 2014, 55, 718–728.
- Wasserman, A.H.; Venkatesan, M.; Aguirre, A. Bioactive Lipid Signaling in Cardiovascular Disease, Development, and Regeneration. Cells 2020, 9, 1391.
- Brezinski, A.D.; Nesto, R.W.; Serhan, C.N. Angioplasty triggers intracoronary leukotrienes and lipoxin A4. Impact of aspirin therapy. Circulation 1992, 86, 56–63.
- Shen, J.; Herderick, E.; Cornhill, J.F.; Zsigmond, E.; Kim, H.S.; Kühn, H.; Guevara, N.V.; Chan, L. Macrophage-mediated 15-lipoxygenase expression protects against atherosclerosis development. J. Clin. Investig. 1996, 98, 2201–2208.
- Serhan, C.N.; Jain, A.; Marleau, S.; Clish, C.; Kantarci, A.; Behbehani, B.; Colgan, S.P.; Stahl, G.; Merched, A.; Petasis, N.; et al. Reduced Inflammation and Tissue Damage in Transgenic Rabbits Overexpressing 15-Lipoxygenase and Endogenous Anti-inflammatory Lipid Mediators. J. Immunol. 2003, 171, 6856–6865.
- Cyrus, T.; Witztum, J.L.; Rader, D.J.; Tangirala, R.; Fazio, S.; Linton, M.F.; Funk, C.D. Disruption of the 12/15-lipoxygenase gene diminishes atherosclerosis in apo E–deficient mice. J. Clin. Investig. 1999, 103, 1597–1604.
- Hasturk, H.; Abdallah, R.; Kantarci, A.; Nguyen, D.; Giordano, N.; Hamilton, J.; Van Dyke, T.E. Resolvin E1 (RvE1) Attenuates Atherosclerotic Plaque Formation in Diet and Inflammation-Induced Atherogenesis. Arter. Thromb. Vasc. Biol. 2015, 35, 1123–1133.
- Virmani, R.; Burke, A.P.; Farb, A.; Kolodgie, F.D. Pathology of the Vulnerable Plaque. J. Am. Coll. Cardiol. 2006, 47, C13–C18.
- Schrijvers, D.M.; De Meyer, G.R.; Kockx, M.M.; Herman, A.G.; Martinet, W. Phagocytosis of Apoptotic Cells by Macrophages Is Impaired in Atherosclerosis. Arter. Thromb. Vasc. Biol. 2005, 25, 1256–1261.
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2009, 10, 36–46.
- Fredman, G.; Hellmann, J.; Proto, J.D.; Kuriakose, G.; Colas, R.A.; Dorweiler, B.; Connolly, E.S.; Solomon, R.; Jones, D.M.; Heyer, E.J.; et al. An imbalance between specialized pro-resolving lipid mediators and pro-inflammatory leukotrienes promotes instability of atherosclerotic plaques. Nat. Commun. 2016, 7, 12859.
- Viola, J.R.; Lemnitzer, P.; Jansen, Y.; Csaba, G.; Winter, C.; Neideck, C.; Silvestre-Roig, C.; Dittmar, G.; Döring, Y.; Drechsler, M.; et al. Resolving Lipid Mediators Maresin 1 and Resolvin D2 Prevent Atheroprogression in Mice. Circ. Res. 2016, 119, 1030–1038.
- Petri, M.H.; Laguna-Fernandez, A.; Arnardottir, H.; E Wheelock, C.; Perretti, M.; Hansson, G.K.; Bäck, M. Aspirin-triggered lipoxin A4 inhibits atherosclerosis progression in apolipoprotein E−/−mice. Br. J. Pharmacol. 2017, 174, 4043–4054.
- Wang, M.; Liu, M.; Zhang, J.; Liu, J.; Ye, J.; Xu, Y.; Wang, Z.; Ye, D.; Zhao, M.; Wan, M. Resolvin D1 protects against sepsis-induced cardiac injury in mice. Biofactors 2020, 46, 766–776.
- Kain, V.; Ingle, K.A.; Colas, R.A.; Dalli, J.; Prabhu, S.D.; Serhan, C.N.; Joshi, M.D.; Halade, G.V. Resolvin D1 activates the inflammation resolving response at splenic and ventricular site following myocardial infarction leading to improved ventricular function. J. Mol. Cell. Cardiol. 2015, 84, 24–35.
- Keyes, K.T.; Ye, Y.; Lin, Y.; Zhang, C.; Perez-Polo, J.R.; Gjorstrup, P.; Birnbaum, Y. Resolvin E1 protects the rat heart against reperfusion injury. Am. J. Physiol. Circ. Physiol. 2010, 299, H153–H164.
- Duffield, J.S.; Hong, S.; Vaidya, V.S.; Lu, Y.; Fredman, G.; Serhan, C.N.; Bonventre, J.V. Resolvin D Series and Protectin D1 Mitigate Acute Kidney Injury. J. Immunol. 2006, 177, 5902–5911.
- Kasuga, K.; Yang, R.; Porter, T.F.; Agrawal, N.; Petasis, N.; Irimia, D.; Toner, M.; Serhan, C.N. Rapid Appearance of Resolvin Precursors in Inflammatory Exudates: Novel Mechanisms in Resolution. J. Immunol. 2008, 181, 8677–8687.
- Foley, J.H.; Conway, E.M. Cross Talk Pathways Between Coagulation and Inflammation. Circ. Res. 2016, 118, 1392–1408.
- Gerdes, N.; Seijkens, T.; Lievens, D.; Kuijpers, M.J.E.; Winkels, H.; Projahn, D.; Hartwig, H.; Beckers, L.; Megens, R.T.A.; Boon, L.; et al. Platelet CD40 Exacerbates Atherosclerosis by Transcellular Activation of Endothelial Cells and Leukocytes. Arter. Thromb. Vasc. Biol. 2016, 36, 482–490.
- Lippi, G.; Montagnana, M.; Salvagno, G.L.; Cicorella, N.; Degan, M.; Minuz, P.; Lechi, C.; Guidi, G.C. Risk stratification of patients with acute myocardial infarction by quantification of circulating monocyte-platelet aggregates. Int. J. Cardiol. 2007, 115, 101–102.
- Chiang, N.; Bermudez, E.A.; Ridker, P.M.; Hurwitz, S.; Serhan, C.N. Aspirin triggers antiinflammatory 15-epi-lipoxin A4 and inhibits thromboxane in a randomized human trial. Proc. Natl. Acad. Sci. USA 2004, 101, 15178–15183.
- Dona, M.; Fredman, G.; Schwab, J.M.; Chiang, N.; Arita, M.; Goodarzi, A.; Cheng, G.; Von Andrian, U.H.; Serhan, C.N. Resolvin E1, an EPA-derived mediator in whole blood, selectively counterregulates leukocytes and platelets. Blood 2008, 112, 848–855.
- Abdulnour, R.-E.E.; Dalli, J.; Colby, J.K.; Krishnamoorthy, N.; Timmons, J.Y.; Tan, S.H.; Colas, R.A.; Petasis, N.; Serhan, C.N.; Levy, B.D. Maresin 1 biosynthesis during platelet–neutrophil interactions is organ-protective. Proc. Natl. Acad. Sci. USA 2014, 111, 16526–16531.
- Norris, P.C.; Arnardottir, H.; Sanger, J.M.; Fichtner, D.; Keyes, G.S.; Serhan, C.N. Resolvin D3 multi-level proresolving actions are host protective during infection. Prostaglandins Leukot. Essent. Fat. Acids 2018, 138, 81–89.
- Bayman, E.; Drake, A.J.; Piyasena, C. Prematurity and programming of cardiovascular disease risk: A future challenge for public health? Arch. Dis. Child. Fetal Neonatal Ed. 2014, 99, F510–F514.
- Kerkhof, G.F.; Breukhoven, P.E.; Leunissen, R.W.; Willemsen, R.H.; Hokken-Koelega, A.C. Does Preterm Birth Influence Cardiovascular Risk in Early Adulthood? J. Pediatr. 2012, 161, 390–396.e1.
- Bassareo, P.P.; Fanos, V.; Puddu, M.; Demuru, P.; Cadeddu, F.; Balzarini, M.; Mercuro, G. Reduced brachial flow-mediated vasodilation in young adult ex extremely low birth weight preterm: A condition predictive of increased cardiovascular risk? J. Matern. Neonatal Med. 2010, 23, 121–124.
- Lazdam, M.; De La Horra, A.; Pitcher, A.; Mannie, Z.; Diesch, J.; Trevitt, C.; Kylintireas, I.; Contractor, H.; Singhal, A.; Lucas, A.; et al. Elevated Blood Pressure in Offspring Born Premature to Hypertensive Pregnancy. Hypertension 2010, 56, 159–165.
- Khodaghalian, B.; Subhedar, N.V.; Chikermane, A. Prostaglandin E2 in a preterm infant with coarctation of the aorta. BMJ Case Rep. 2019, 12, e230910.
- Desai, J.; Aggarwal, S.; Lipshultz, S.; Agarwal, P.; Yigazu, P.; Patel, R.; Seals, S.; Natarajan, G. Surgical Interventions in Infants Born Preterm with Congenital Heart Defects: An Analysis of the Kids’ Inpatient Database. J. Pediatr. 2017, 191, 103–109.e4.
- Callahan, P.; Quivers, E.; Bradley, L.; Sell, J.; Martin, G. Echocardiographic Evidence for a Ductal Tissue Sling Causing Discrete Coarctation of the Aorta in the Neonate: Case Report. Pediatr. Cardiol. 1998, 19, 182–184.
- Heymann, M.A.; Rudolph, A.M. Control of the ductus arteriosus. Physiol. Rev. 1975, 55, 62–78.
- Hung, Y.-C.; Yeh, J.-L.; Hsu, J.-H. Molecular Mechanisms for Regulating Postnatal Ductus Arteriosus Closure. Int. J. Mol. Sci. 2018, 19, 1861.
- Noori, S.; McCoy, M.; Friedlich, P.; Bright, B.; Gottipati, V.; Seri, I.; Sekar, K. Failure of Ductus Arteriosus Closure Is Associated With Increased Mortality in Preterm Infants. Pediatrics 2009, 123, e138–e144.
- Revermann, M. Pharmacological inhibition of the soluble epoxide hydrolase—from mouse to man. Curr. Opin. Pharmacol. 2010, 10, 173–178.
- Imig, J.D.; Hammock, B.D. Soluble epoxide hydrolase as a therapeutic target for cardiovascular diseases. Nat. Rev. Drug Discov. 2009, 8, 794–805.
- Morgan, L.A.; Olzinski, A.R.; Upson, J.J.; Zhao, S.; Wang, T.; Eisennagel, S.H.; Hoang, B.; Tunstead, J.R.; Marino, J.P.; Willette, R.N.; et al. Soluble Epoxide Hydrolase Inhibition Does Not Prevent Cardiac Remodeling and Dysfunction After Aortic Constriction in Rats and Mice. J. Cardiovasc. Pharmacol. 2013, 61, 291–301.
- Serhan, C.N.; Chiang, N.; Dalli, J. The resolution code of acute inflammation: Novel pro-resolving lipid mediators in resolution. Semin. Immunol. 2015, 27, 200–215.
- Jannaway, M.; Torrens, C.; A Warner, J.; Sampson, A.P. Resolvin E1, resolvin D1 and resolvin D2 inhibit constriction of rat thoracic aorta and human pulmonary artery induced by the thromboxane mimetic U46619. Br. J. Pharmacol. 2018, 175, 1100–1108.
- Colas, R.A.; Souza, P.R.; Walker, M.E.; Burton, M.; Zaslona, Z.; Curtis, A.M.; Marques, R.M.; Dalli, J. Impaired Production and Diurnal Regulation of Vascular RvD n-3 DPA Increase Systemic Inflammation and Cardiovascular Disease. Circ. Res. 2018, 122, 855–863.


