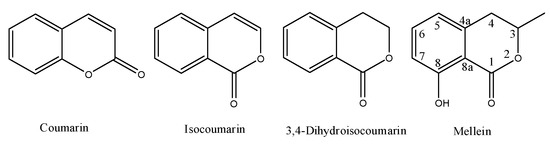
2.1. Melleins from Fungi
Fungi are the most important source of melleins, and (R)-(-)-mellein (1, ) is the most common among this group.
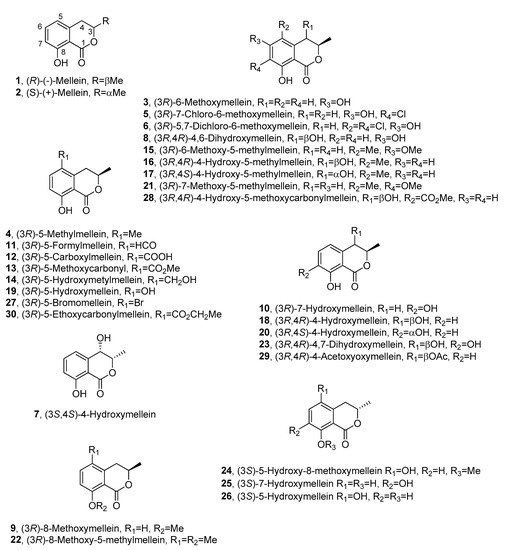
Figure 2. The structures of 1–30.
Compound
1 was isolated for the first time in 1933 from the fungus
Aspergillus melleus [3] and named ocracin, as cited above. However, its structure was determined only in 1955
[11] and the
R absolute configuration (AC) at C-3 was successively assigned
[12][13]. Its enantiomer, the (
S)-(+)-mellein (
2, ), is also known as a natural product, but it is produced by few species of fungi compared with
1. In particular, compound
2 was firstly isolated from an unidentified fungus
[14] and then from the cultures of the insect pathogen
Fusarium larvarum, together with five other secondary metabolites
[15].
A structural analogue of
1 and
2 was isolated from
Sporormia bipartis and characterized as 6-methoxymellein (
3, )
[16]; another analogue was isolated from the plant pathogen
Fusicoccum amygdali, together with the main phytotoxic metabolite fusicoccin A. The compound, whose structure was elucidated by NMR, was found to be 5-methylmellein (
4, ). In this study,
4 inhibited conidia germination in some fungi, but it had no detectable phytotoxicity in vitro
[17]. Compound
3 was obtained as the main metabolite produced by the fungus
Sporormia affinis together with two chlorinated analogues, which were characterized as 7-chloro-6-methoxymellein (
5, ) and 5,7-dichloro-6-methoxymellein (
6, ) by NMR and mass spectra
[18].
Studies carried out to evaluate the production of mycotoxins by
Aspergillus oniki 1784, allowed to isolate
1 together with two related compounds, which were characterized as 3-methyl-4,8-dihydroxy-3,4-dihydroisocoumarin and 3-methyl-3,8-dihydroxy-3,4-dihydroisocoumarin. Their LD
50 values for mice were measured by intraperitoneal injection, but their stereochemistry was not determined
[19]. Successively, compound
1 together with
cis-4-hydroxymellein and other five metabolites were isolated from
Lasidiplodia theobromae culture filtrates, which inhibited the green plant growth. However, for
1 and its analogues, no biological activity was reported
[20]. One of the stereoisomers of 4-hydroxymellein was also isolated together with
1 from
Aspergillus ochraceus [21][22]. The AC of one of the two enantiomers of
cis-4-hydroxymellein was determined when it was purified from the mycelium of
Cercospora taiwanensis, together with
2, and characterized as
cis-(3
S,4
S)-4-hydroxymellein (
7, )
[23]. Furthermore, a 4,6-dihydroxymellein (
8, ) was isolated in a screening of 61 other species of
Cercospora [24].
(
R)-(-)-mellein (
1), (
R)-8-methoxymellein (
9, ) and (3
R)-7-hydroxymellein (
10, ) were isolated from
Septoria nodorum by Devys and co-authors and characterized by NMR spectroscopy
[25].
A series of dihydroisocoumarin derivatives were isolated in a systematic study on metabolites of the Xylariaceous fungi belonging to
Hypoxylon and
Numularia species. 5-methylmellein (
4) was produced by almost all species studied, whereas
1 was produced by
Hypoxylon fragiforme, Hypoxylon howeianum, Hypoxylon haematostroma, Hypoxylon venusfuissimum and
Hypoxylon deustum. In the same study, five other analogues of mellein were isolated and identified as 5-formyl- (
11, ), 5-carboxy- (
12, ), 5-methoxycarbonyl (
13, ), 5-hydroxymethyl (
14, ), and 6-methoxy-5-methyl- (
15, ) mellein.
11 was produced by
Numularia discreta and
Numularia broomiana, 12 by
Hypoxylon mammatum, Hypoxylon illitum and
N. discreta, 13 by
H. mammatum, 14 by
H. illitum and
15 by
Hypoxylon atropunctatum [26].
Mellein 5-carboxylic acid (
12), together with 5-methyl mellein (
4), (
S)-(+)-mellein (
2) and other compounds, was isolated from
Phomopsis oblonga in a study for the production of elm bark beetle boring and feeding deterrents. Compound
2 was obtained as minor metabolite, while
4 and
12 were active against adult females of
Scolytus sp. beetles
[27]. The AC at C-3 of (-)-5-carboxylmellein (
12) and (-)-5-hydroxylmethylmellein (
15) was assigned as
R by chemical correlations when they were isolated from
Valsa ceratosperma, a fungus inducing apple canker. The same fungus also synthesizes (-)-5-methylmellein (
4), and two new compounds which were characterized as
cis-(3
R,4
R)-(-)-4-hydroxy-5-methylmellein (
16, ) and
trans-(3
R,4
S)-(+)-4-hydroxy-5-methylmellein (
17, ). All the five compounds were phytotoxic to apple shoots and lettuce seedlings
[28].
The first report of
1 in the genus
Nectria occurred in 1986 when it was isolated from
Nectria fuckeliana by Ayer and Shewchuk
[29].
Five phytotoxic metabolites were purified from the culture filtrates of
Botryosphearia obtusa (Schw.) Shoemaker, the causal agent of frogeye leaf spot and fruit black rot of apple. Among them, (
R)-(-)-mellein (
1),
cis-(3
R,4
R)-(-)-4-hydroxymellein (
18, ) and (3
R)-5-hydroxymellein (
19, ) were identifiedusing spectroscopic and optical methods
[30].
19 was isolated for the first time as a fungal metabolite and the phytotoxicity of all metabolites was tested by leaf puncture assay on seventeen apple cultivars and eight weed species. The apple cultivars Supergold and Silverspure and the weeds species prickely sida and morning glory were the most sensitive species to all the phytotoxins
[31]. Successively, (
R)-(-)-mellein (
1) was isolated as a phytotoxic metabolite, together with citrinin, tyrosol and α–acetylorcinol, from
Stagonospora apocynin, which caused leaf spot disease on hemp dogbane (
Apocynum cannabinum L.). All the compounds were non–specific toxins causing necrosis when assayed on hemp dogbane leaves, and those of eight other weed species
[32].
In a study carried out to select in vitro wheat embryos with a high level of resistance,
1 was identified in the fungal organic extract of
Septoria nodorum (Berk.)
[33]. (3
R,4
S)-(-)-4-hydroxymellein (
20, ), together with (3
R,4
R)-(-)-4-hydroxymellein (
18), was isolated for the first time from the same organic extract
[34].
Phoma tracheiphila, the fungus responsible for “mal secco” citrus disease, also produced (
R)-(-)-mellein (
1). Compound
1 was phytotoxic when tested at a concentration of 100 mg/mL in tomato cuttings and caused 100% mortality of
Artemia salina larvae at 200 mg/mL
[35].
Compound
1 was isolated together with other four compounds in a study conducted on 85
Pezicula strains isolated as endophytes from living branches of ten deciduous and coniferous trees. All the compounds demonstrated strong fungicidal and herbicidal activity, and to a lesser extent, algicidal and antibacterial activity. Their production was taxonomically significant
[36].
(
R)-(-)-mellein (
1), (3
R,4
R)-hydroxymellein (
18) and (3
R,4
S)-hydroxymellein were also isolated from
Microsphaeropsis sp. in a study conducted on some fungi obtained from marine sponges. In the same study (3
R)-6-methoxymellein (
3) and (3
R)-6-methoxy-7-chloromellein (
5) were also isolated from the culture extracts of the
Coniothyrium sp. All these mellein analogues showed antifungal activity against
Eurotium repens and
Ustilago violacea [37].
A crude extract of
Aspergillus ochraceus inhibited the final stage of hepatitis C virus (HCV) replication. A bio-guided purification of the extract afforded the known (
R)-(-)-mellein (
1), together with circumdatins G and F, which were identified by NMR spectroscopy. Compound
1 inhibited HCV protease with an IC
50 value of 35 μM
[38].
Two novel melleins, namely, 7-methoxy-5-methylmellein (
21, ) and 8-methoxy-5-methylmellein (
22, ), together with three known dihydroisocoumarins and a tetralone derivative, were isolated from a culture filtrate of
Cytospora eucalypticola. Their structures were elucidated by spectroscopic methods. Compounds
21 and
22 showed moderate antifungal and antibacterial activity against Gram-positive bacteria
[39].
Apiospora montagnei, a marine fungus, isolated from the North Sea alga
Polysiphonia violacea, produced a plethora of secondary metabolites comprising (
R)-(-)-mellein (
1) and (
R)-8-methoxymellein (
9)
[40]. Two novel compounds, namely, the monomethyl ester of 9-hydroxyhexylitaconic acid and the (-)-enantiomer of the known (+)-hexylitaconic acid, were also isolated and characterized by spectroscopic methods. The crude fungal extract and all the metabolites did not show antibacterial, antifungal and antialgal activity
[40].
Sphaeropsis sapinea was isolated from declining pine (
Pinus radiata) plants in Sardinia and studied for its ability to produce phytotoxic metabolites
[41].
S. sapinea was grown in liquid culture and the purification of the corresponding organic extract afforded the three already known (
R)-(-)-mellein (
1), (3
R,4
R)-4-hydroxymellein (
18) and (3
R,4
S)-4-hydroxymellein (
20) isolated for the first time from this fungus. When assayed for phytotoxic and antifungal activities on host and non-host plants and on some phytopathogenic fungi,
1 was the most active compound, while
18 and
20 showed only a synergic effect in both tests
[41]. The same melleins were isolated from
Botryosphaeria mamane PSU-M76, along with other three known secondary metabolites and a dihydrobenzofuran derivative named botryomaman
[42]. Their antibacterial activity against
Staphylococcus aureus ATCC 25923,
S. aureus SK1 and compounds
1,
18 and
20 was tested; they were inactive with equal MIC values of > 128 μg/mL
[42].
Among a collection of 250 foliar endophytes of
Picea glauca, several isolates produced toxic metabolites against
Choristoneura fumiferana (spruce budworm)
[43]. Three of them (strains CBS 120381, CBS 120379 and CBS 120380) were selected for isolation and characterization of phytotoxic metabolites by LC-MS and LC-NMR. The DNA sequencing revealed that CBS 120381 belongs to the Xylariaceae and it was near but not identical to
Nemania serpens. The sequencing data indicated that CBS 120379 and CBS 120380 were both species of
Lophodermium, with CBS 120379 most like fungi in the
Rhytistimataceae. Two of them produced at least one mellein. CBS 120379 synthesized (
R)-(-)-mellein (
1), while CBS 120381 produced three known melleins: 5-methyl- (
4), 5-formyl- (
11) and 5-methoxycarbonyl- (
13) mellein
[43].
Botryosphaeria obtusa, the teleomorph of
Diplodia seriata, is a pathogen associated with grapevine Botryosphaeria dieback. The fungus was studied for its ability to synthesize phytotoxic metabolites, which were purified and characterized by spectroscopic data
[44]. The three known melleins, (
R)-(-)-mellein (
1), (3
R)-7-hydroxymellein (
10) and (3
R,4
R)-4-hydroxymellein (
18), and a novel mellein characterized as 4,7-dihydroxymellein (
23, ) were isolated from the liquid culture of this fungus
[44]. When assayed on grapevine leaves,
23 was the most active compound, inducing full leaf necrosis with minimum inhibitory concentrations (MIC) of 2 μg/mL
[44]. Other Botryosphaeriaceae, such as
Bothryosphaeria rhodina PSU-M35 and PSU-M114, were studied for secondary metabolite production because they showed antibacterial activity against
S. aureus, both standard ATCC 25922 (SA) and methicillin-resistant (MRSA) strains. (
R)-(-)-mellein (
1), (3
R,4
R)-4-hydroxymellein (
18), (3
R,4
S)-4-hydroxymellein (
20) and (3
R)-5-hydroxymellein (
19) were recognized by spectroscopic and physic properties. Compounds
1,
18,
19 and
20 showed good activity against both strains; however, the most active compound isolated was (3
S)-lasiodiplodin
[45].
Tubercularia sp. TF5 was isolated from the bark of
Taxus mairei collected in Fujian Province, southeast China. Taxol, the well-known anticancer compound, was produced by this strain. Thus, it was studied for the production of other bioactive metabolites. The chromatographic purification of the culture filtrate extract yielded 5-carboxymellein (
12) which was identified by spectroscopic data
[46].
Neofusicoccum parvum is the one of the most virulent Botryosphaeriaceae species causing heavy grapevine trunk diseases. (3
R,4
R)-4-hydroxymellein (
18), (3
R,4
S)-4-hydroxymellein (
20), isosclerone and tyrosol were produced by this fungus and identified by spectroscopic methods. All the compounds were tested to evaluate their phytotoxic activities on tomato cuttings at different concentrations, and they showed toxicity ranging from slight to drastic leaf wilting, and compound
18 showed the highest phytotoxicity
[47].
Diplodia africana, the causal agent of branch dieback on
Juniperus phoenicea, belonging to the same fungal family of the Botryosphaeriaceae, produced phototoxic metabolites
[48]. Two novel phytotoxic dihydrofuropyran-2-ones, named afritoxinones A and B, were isolated, together with known compounds, from the culture filtrates of
D. africana. Besides afritoxinones A and B, the three well-known melleins (
R)-(-)-mellein (
1), (3
R,4
R)-4-hydroxymellein (
18) and (3
R,4
S)-4-hydroxymellein (
20) were isolated
[48].
Endophytic fungi living in the intracellular spaces of plants are able to synthesize novel bioactive secondary metabolites. Indeed, the endophytic fungal strain BCRC 33717, obtained from the bark of the medicinal plant
Cinnamomum sp., produced a plethora of secondary metabolites, including four melleins, which were recognized as the 5-formyl- (
11), 5-carboxy- (
12) and 5-hydroxy- (
19) mellein, and the novel (3
S)-5-hydroxy-8-
O-methylmellein (
24, )
[49]. Their structures were determined by NMR and MS methods
[49].
Annulohypoxylon squamulosum BCRC 34022, obtained from the same medicinal plants, when grown on a long-grain rice fermented culture, synthesized eleven different metabolites, which were extracted with
n-BuOH-soluble 95% EtOH. Among the secondary bioactive metabolites purified from these extracts, three melleins were characterized by spectroscopic methods as(+)-(
S)-mellein (
2), (
S)-7-hydroxymellein (
25, ), and (
S)-5-hydroxymellein (
26, ).
[50]. Compounds
25 and
26 were isolated for the first time and are the enantiomers of the previously reported compounds
10 and
19. When assayed on MCF-7 (human breast adenocarcinoma), NCIH460 (non-small-cell lung cancer) and SF-268 (glioblastoma cells), compound
2 possessed moderate toxicity against MCF-7, NCI-H460 and SF-268 cancer cell lines, while
25 and
26 showed weak to negligible activities against all the three cell lines
[50].
To keep talking about endophytic fungi,
Epicoccum nigrum, an ascomycete fungus distributed worldwide, colonizes different types of soils and host plants, and was used as a biocontrol agent for plant pathogens.
E. nigrum wild type P16 produced secondary metabolites, including (
R)-(-)-mellein (
1)
[51]. Three
E. nigrum agro-transformants, namely, P16–17, P16–47set and P16–91, were studied in order identify the genes related to the synthesis of a new natural compound produced by
E. nigrum P16. The comparison of the extracts of the wild type and the transformants by GC-MS, revealed that the mutants were capable of producing (
R)-5-hydroxymellein (
19) as well
[51].
Biscogniauxia nummularia (Bull.), a seemingly endophytic fungus isolated from the plum yew
Cephalotaxus harringtonia, produced a new guaiane sesquiterpene together with a previously known terpenoid, xylaranol B, and two mellein derivatives identified as 5-methylmellein (
4) and 8-methoxy-5-methylmellein (
22)
[52].
Seimatosporium sp. was studied for its ability to synthesize biologically active compounds in a program planned to investigate endophytic fungi for new products for pharmacy and plant protection
[53]. (
R)-(-)-Mellein (
1), (3
R,4
R)-4-hydroxymellein (
18), (3
R,4
S)-4-hydroxymellein (
20), (3
R)-5-hydroxymethylmellein (
14) and (3
R,4
S)-4-hydroxy-5-methylmellein (
17) were isolated from fermentation extracts of
Seimatosporium sp. and identified by spectroscopic methods
[53].
Two novel succinic acid derivatives, xylacinic acids A and B, were isolated, along with (
R)-8-methoxymellein (
9), (3
R)-5-methoxycarbonylmellein (
13) and (3
R)-5-hydroxymellein (
19), from the mangrove-derived fungus
Xylaria cubensis PSU-MA34
[54]. Al the metabolites isolated were tested for antibacterial activity against
S. aureus ATCC 25923 and MRSA and for cytotoxicity against KB cells; however, the melleins were inactive
[54].
The extracts from eleven fungal strains associated with
Eudistoma vannamei were assayed against two cell lines asthe colon and melanoma cancer cell lines
[55].
Aspergillus sp. yielded the most promising extract. Its potato dextrose broth extract was purified by bio-guided chromatographic method, and (
R)-(-)-mellein (
1), (3
R,4
R)-4-hydroxymellein (
18) and (3
R,4
S)-4-hydroxymellein (
20) were isolated, along with penicillic acid. All isolated compounds were tested for their cytotoxicity; however, only penicillic acid showed cytotoxic activity (cell growth inhibitions > 95%)
[55].
The bioactive metabolites were produced by an endophytic fungus, identified as
Nigrospora sp. by rDNA sequence analysis, and four of them were identified by comparison of the
1H-NMR and
13C-NMR spectroscopic data with those previously reported. Among them, (
R)-(-)-mellein (
1) was identified and showed only weak antifungal activity
[56]. In another investigation the crude extracts from cultures of
Pezicula livida were studied for larvicidal activity. The bio-guided chromatographic purification of the extract also afforded (
R)-(-)-mellein (
1), and its larvicidal activity was reported the first time with an LC
50 value of 1.4 ppm against
Aedes aegypti [57].
An interesting study was conducted by Yun and co-authors (2013), who added NaBr to the fermentation of the fungus
Aspergillus ochraceus [58]. This resulted in the production of a new brominated mellein analogue; namely, (3
R)-5-bromomellein (
27, ). Furthermore, three known compounds, (
R)-(-)-mellein (
1), clavatol and circumdatin A, were also recognized in the same culture. The structure of
27 was assigned through spectroscopic data analyses. All the isolated compounds showed mild radical scavenging activity against 1,1-diphenyl-2-picrylhydrazyl (DPPH)
[58].
The organic extract from culture broths of
Arthrinium state of
Apiospora montagnei afforded the main metabolites, which were identified as (
R)-(-)-mellein (
1) and (3
R,4
R)-4-hydroxymellein (
18) according to their spectroscopic and physical data. In addition, their activity against
Schistosoma mansoni (adult worms) was tested. Despite the structural similarity,
1 caused the death of 100% of parasites (both male and female) at 200 μg/mL, whereas compound
18 caused the death of 50% of adult worms at 12.5 μg/mL and 100% at 50 μg/mL
[59].
Xylaria sp. PSU-G12, isolated from
Garcinia hombroniana, when grown in liquid culture, yielded a corresponding organic extract displaying antioxidant activity
[60]. A plethora of secondary metabolites were isolated and characterized by spectroscopic methods, and among them a novel mellein analogue was isolated and characterized as (3
R,4
R)-5-carbomethoxy-4-hydroxymellein, and then named xylarellein (
28, ). Three known melleins were also identified as (3
R)-5-methylmellein (
4), (3
R)-5-carboxylmellein (
12) and (3
R)-5-methoxycarbonilmellein (
13). All the isolated compounds were evaluated for antioxidant activity in DPPH assays, but none of them exhibited antioxidant activity, highlighting the possibility that the antioxidant activity of the crude extract may involve a synergistic effect
[60].
Two formerly undescribed polyketide metabolites were obtained from the cultures of an endophytic fungus isolated from
Meliotus dentatus. The two compounds appeared to be, based on spectroscopic data, a new mellein named
cis-4-acetoxyoxymellein (
29, ) and one of its derivatives
[61]. The two compounds were tested in an agar diffusion assay for their antifungal, antibacterial and algicidal activities against
Botrytis cinerea,
Septoria tritici,
Phytophthora infestans,
Microbotryum violaceum,
Escherichia coli,
Bacillus megaterium and
Chlorella fusca. Both metabolites displayed strong antibacterial activity, especially towards
Escherichia coli and
Bacillus megaterium. In addition, both displayed algicidal activity against
C. fusca and good antifungal activity against
M. violaceum,
B. cinerea and
S. tritici [61].
The endophyte
Xylaria sp., isolated from a surface-sterilized Concord grape leaf (
Vitis labrusca), grown in liquid culture, produced seven compounds, among which two mellein derivatives were isolated and identified by spectroscopic data as (3
R)-5-methoxycarbonylmellein (
13) and (3
R)-5-carboxylmellein (
12). However, no biological activities were tested for these two metabolites
[62].
(
R)-(-)-mellein (
1) was also produced by
Lasiodiplodia sp. ME4-2, an endophytic fungus obtained from the floral sections of
Viscum coloratum [63], and from
Pezicula sp., an endophyte obtained from twigs of
Forsythia viridissima, Zhejiang Province, Southeast China
[64]. In this latter study, (
R)-(-)-mellein (
1) showed growth inhibition against nine plant pathogenic fungi, especially
Botrytis cinerea and
Fulvia fulva with EC
50 values below 50 μg/mL
[64].
From the solid culture of the endophytic fungus
Xylaria sp. SNB-GTC2501, which was obtained from the leaves of
Bisboecklera microcephala, the two already known mellein derivatives
4 and
12 were obtained, whose antimicrobial potential was assessed against human pathogens (i.e.,
Staphylococcus aureus,
Trichophyton rubrum and
Candida albicans). However, their minimal inhibitory concentrations were more than 128 μg/mL. In addition,
4 and
12 were not cytotoxic towards MRC5 cells (IC
50 >100 μM)
[65].
(3
R)-5-methylmellein (
4), (3
R)-5-formylmellein (
11) and (3
R)-5-carboxymellein (
12), were also synthesized, together with two undescribed dihydronaphthalenones and the known 2,6-dihydroxy-4-methylacetophenone, by the endophytic fungus
Nodulisporium sp., isolated from
Antidesma ghaesembilla. No biological activities were reported for the mellein derivatives
4,
11 and
12
[66].
The endophytic fungus
Botryosphaeria sp. KcF6 derived from the mangrove plant
Kandelia candel produced (3
R)-8-methoxymellein (
9) and (3
R,4
S)-4-hydroxylmellein (
20), which were identified by spectroscopic data
[67]. The isolated compounds were evaluated for their cytotoxic and anti-inflammatory (COX-2) activities, but
9 and
20 were inactive
[67].
The previously undescribed (3
R)-5-ethoxycarbonylmellein (
30, ) was isolated from the fungus
Marasmiellus ramealis isolated in China together with the known
13 and other nine compounds. The structure of the new compound was elucidated by spectroscopic methods, but no activity was reported
[68].
The ability of
N. parvum to produce phytotoxins was further investigated by Abou-Mansour et al.
[69]. (
R)-(-)-mellein (
1), (3
R,4
R)-4-hydroxymellein (
18) and (3
R,4
S)-4-hydroxymellein (
20), already reported as phytotoxic metabolites produced by another strain of
N. parvum [47], were isolated together with 3-hydroxymellein and other compounds belong to different chemical families. 3-Hydroxymellein (
31, ) was previously isolated from
Aspergillus oniki 1784
[19], but this study provided its first
1H and
13C NMR data. Its 3
R AC was assigned by comparing its CD spectrum with that of
1.
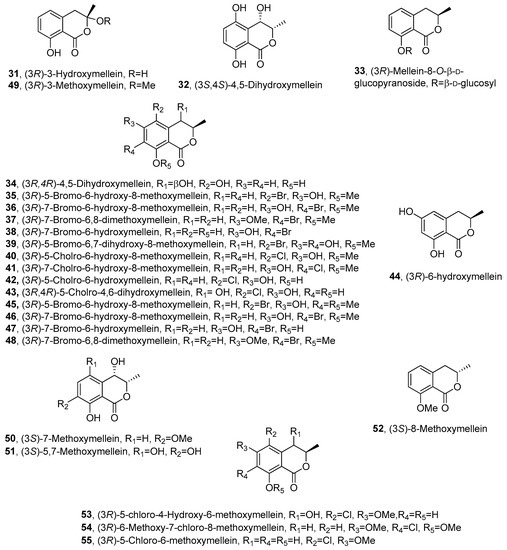
Figure 3. The structures of compounds 31–55.
The phytotoxicity of the main isolated compounds were assessed 48 h after-inoculation at concentrations of 100 and 200 μg/mL, on leaf discs of
Vitis vinifera cv. Chardonnay. All the compounds tested induced necrosis on host plant leaves, and among the four melleins, (3
R)-3-hydroxymellein (
31) was the most active at the lower concentration
[69].
Six species of
Lasiodiplodia isolated in Brazil and causing Botryosphaeria dieback of grapevines were studied to evaluate their production of phytotoxic secondary metabolites. Some mellein derivatives were produced, as ascertained by LC/MS. In particular,
Lasiodiplodia brasiliense synthesized (3
R,4
S)-4-hydroxymellein (
20), and
Lasiodiplodia euphorbicola produced (
R)-(-)-mellein (
1), (3
R,4
R)- and (3
R,4
S)- 4-hydroxymellein (
18 and
20)
[70].
The production of
1 by the endophyte
Lasiodiplodia theobromae was confirmed in a study on the metabolomics-guided isolation of anti-trypanosomal metabolites. However,
1 was inactive when tested against
Trypanosoma brucei brucei [71]. Compound
1 was also isolated together tyrosol and a new isochromanone, named fraxitoxin, from liquid cultures of
Diplodia fraxini, a pathogen involved in the etiology of canker and dieback disease of
Fraxinus spp. in Europe
[72].
Paraconiothyrium sporulosum YK-03, a marine-derived fungus, also produced
1 together with seven mellein derivatives, including the previously undescribed (3
S,4
S)-4,5-dihydroxymellein (
32, ) and (
R)-(-)-mellein-8-
O-β-D-glucopyranoside (
33, ). Their structures and ACs were determined by comprehensive spectroscopic and computational electronic circular dichroism (ECD). Furthermore, their monosaccharide composition was determined by HPLC coupled with optical rotation detector. The known compounds were identified by spectroscopic methods as (3
R)-7-hydroxymellein (
10), (3
R)-5-hydroxymellein (
19), (3
R,4
R)-4-hydroxymellein (
18), (3
R,4
S)-4-hydroxymellein (
20) and (3
R,4
S)-4,5-dihydroxymellein (
34, )
[73]. Compound
34 was previously isolated from the marine fungus
Phomopsis sp. (number ZH-111)
[73]. All the compounds were assessed for their cytotoxic activities against the human cancer cell lines A549 and MCF-7. However, none of them showed meaningful cytotoxicity against these two cell lines
[74].
The endophytic fungus
Lachnum palmae, obtained from
Przewalskia tangutica, was exposed to a histone deacetylase inhibitor SAHA and produced seven previously undescribed halogenated dihydroisocoumarins, named palmaerones A-G (
35-
41, ), together with eleven known dihydroisocoumarins. They were identified by spectroscopic and optical methods as
1,
3,
18,
19,
20, (3
R)-5-bromo-6-hydroxy-8-methoxymellein (
35), (
3R)-7-bromo-6-hydroxy-8-methoxymellein (
36), (3
R)-7-bromo-6,8-dimethoxymellein (
37), (3
R)-7-bromo-6-hydroxy-mellein (
38), (3
R)-5-bromo-6,7-dihydroxy-8-methoxy-mellein (
39), (3
R)-5-chloro-6-hydroxy-8-methoxy-mellein (
40), (3
R)-7-chloro-6-hydroxy-8-methoxy-mellein (
41), (3
R)-5-cholro-6-hydroxymellein (
42, ), (3
R,4
R)-5-cholro-4,6-dihydroxymellein (
43, ), (3
R)-6-hydroxymellein (
44, ), (3
R)-5-bromo-6-hydroxy-8-methoxy-mellein, named palmaerone A (
45, ), (3
R)-7-bromo-6-hydroxy-8-methoxy-mellein, named palmaerone B (
46, ) and (3
R)-7-bromo-6-hydroxy-mellein, named palmaerone D (
47, )
[75]. The three melleins
45-
47 were previously isolated from the same fungus together with the other mellein derivative, namely, (3
R)-7-bromo-6,8-dimethoxy-mellein, which was named palmerin C (
48, )
[76]. Compounds
35-
41 showed antimicrobial activities against the strains of
Candida neoformans,
Penicillium sp.,
Candida albicans,
Bacilus subtilis and
S. aureus, and compound
39 showed potential antimicrobial activity against all the test strains with the MIC value in the range of 10–55 μg/mL. Metabolites
35 and
39 exhibited mild inhibitory effects on nitric oxide (NO) production in lipopolysaccharide (LPS)-induced RAW 264.7 cells, with the IC
50 values of 26.3 and 38.7 μM, respectively and no evident toxicities were observed at 50 μM. Compound
39 showed weak cytotoxicity against HepG2 with the IC
50 value of 42.8 mM
[76].
An interesting study was carried out on 579 fungal culture extracts to identify the bioactive compounds capable of inhibiting histone deacetylase activity of Sirtuin A (SirA), produced by the fungus
Aspergillus nidulans [77]. Sirtuin is a family of histones that are implicated in fungal growth and secondary metabolite production. Eight fungal strains, belonging to different families, showed to produced SirA inhibitors, and among them,
Didymobotryum rigidum JCM 8837 showed to produce 5-methylmellein (
4). This mellein derivative was found to inhibit SirA activity with IC
50 of 120 μM. In addition, in another experiment compound
4 was added to
A. nidulans cultures. This resulted in an increased secondary metabolite production. In conclusion, the results highlighted that 5-methylmellein (
4) can modulate fungal secondary metabolism and is a potential tool for screening novel compounds derived from fungi
[77].
Compound
1 was isolated from the culture filtrates of
Diplodia mutila FF18
Diplodia seriata H141a,
Neofusicoccum australe VP13 and
Neofusicoccum luteum, as resulted from a recent study on phytotoxic metabolites produced by nine species of Botryosphaeriaceae involved in grapevine dieback in Australia. From
N. luteum, 18 and
20 were also isolated
[78].
Sardiniella urbana, a pathogen of European hackberry trees in Italy, was investigated for its ability to produce secondary metabolites. It produced
1,
18 and
20, which were identified by spectroscopic methods. These compounds were assayed for their phytotoxic, antifungal and zootoxic activities, and among them, only (
R)-(-)-mellein was found to be active. In particular,
1 displayed from significant to weak activity towards all plant pathogens tested at 0.2 mg/plug.
Athelia rolfsii,
Botrytis cinerea and
Sclerotinia sclerotiorum were the most sensitive species. On the contrary,
Alternaria brassicicola,
Fusarium graminearum and
Phytophthora cambivora were less sensitive. In the
Artemia salina bioassay,
1 caused 100% larval mortality at 200 μg/mL. The LC
50 value was 102 μg/mL after 36 h of exposure to the metabolite
[79].
Aspergillus flocculus, an endophyte isolated from the stem of the medicinal plant
Markhamia platycalyx, was investigated for its ability to synthesize bioactive anticancer and anti-trypanosome secondary metabolites. From the fermentation culture of the fungus were isolated several metabolites belonging to different classes of natural compounds. Among them were isolated some mellein derivatives identified as
1, 18,
19,
20,
31,
34 and botryoisocoumarin A (
49, ). Compounds
18,
34,
49 and
1 inhibited the growth of chronic myelogenous leukemia cell line K562 at 30 μM. Compound
31 exhibited an inhibition of 56% to the sleeping-sickness-causing parasite
Trypanosoma brucei brucei [80]. Compound
49, characterized as (3
R)-3-methoxymellein, was also previously obtained from the fermentation culture of
Botryosphaeria sp. F00741, isolated from the plant epidermis of
Avicennia marina [81].
Compound
1 was also recently isolated from the endophytic fungus
Colletotrichum gloeosporioides GT-7, obtained from
Uncaria rhynchophylla [82].
Xylaria sp. SWUF09-62, a Basidiomycota fungus belonging to the Xylariaceae family, was investigated to explore its ability to produce natural products with anti-inflammation and anti-proliferation activities
[83]. This research led to the isolation of several melleins derivatives which were identified by spectroscopic methods as (3
S)-7-methoxymellein (
50, ) and (3
S)-5,7-dihydroxymellein (
51, ), and their ACs were determined by ECD experiments. (3
S)-8-methoxymellein (
52, ), previously synthesized by Kerti et al.
[84], was isolated for the first time as a natural compound. In addition, (
S)-(+)-mellein (
2), (3
R)-5-methoxycarbonylmellein (
13), (3
R,4
R)-4-hydroxymellein (
18), (3
R,4
S)-4-hydroxymellein (
20) and (3
S)-7-hydroxymellein (
25), were also isolated. Anti-inflammatory activity screening was carried out by measuring the reduction of NO production in LPS-induced RAW264.7 macrophage cells, and the mellein derivatives showed different degrees of activity. Compound
51 exhibited anti-inflammatory properties by reducing nitric oxide production in LPS-stimulated RAW264.7 cells, indicating possible chemo-preventative and chemo-therapeutic properties
[83].
The study conducted by Inose et al.
[85] demonstrated the potential of density functional theory (DFT)-based calculations and ECD spectral calculations for structural elucidation of natural compounds. The extract of
Periconia macrospinosa KT3863 was studied for secondary metabolite production and two new chlorinated melleins, (3
R)-5-chloro-4-hydroxy-6-methoxymellein and (3
R)-7-chloro-6-methoxy-8-methoxymellein (
53 and
54, ), were isolated. Furthermore, the authors reported for the first time the complete characterization of the physical properties of the previously isolated (3
R)-5-chloro-6-methoxymellein (
55, )
[86]. The results of (DFT)-based calculations were used to estimate the values of the
13C chemical shifts and the spin coupling constants and compare them with experimental data collected by HMBC experiment. The calculations allowed them to determine the relative configurations of
53. In addition, the ACs of
53–55 were established by comparing the experimental ECD spectra with those obtained by time-dependent DFT calculations. The data showed that
53 afforded an ECD spectrum that was almost the mirror image of that of
54. Finally, the authors studied the antifungal activities of
53–55 with
Cochliobolus miyabeanus as the model organism; unfortunately no significant inhibition was observed
[85].
 +1 point
+1 point