Cultivated cardoon (Cynara cardunculus var. altilis L.) is a promising candidate species for the development of plant cell cultures (PCC) suitable for large-scale biomass production and recovery of nutraceuticals. We set up the first successful protocol for the stable genetic transformation via Agrobacterium tumefaciens of cardoon PCC, and the heterologous overexpression of a MYB transcription factor from Arabidopsis has proven a valuable strategy for the improvement of their biorefinery potential.
1. Introduction
Among the family of Asteraceae,
Cynara cardunculus L. (2n = 2x = 34) is a species of perennial herbaceous plants with annual growth cycle; the species consists of three closely related botanical varieties: the wild cardoon (var.
sylvestris), the globe artichoke (var.
scolymus), and the cultivated cardoon (var.
altilis)
[1]. Given the full cross-compatibility between these varieties, in addition to phenotypic and genetic evidences, previous studies concluded that both the cultivated cardoon and the globe artichoke were domesticated from wild cardoons
[2][3]. Cultivated cardoons are well-adapted to the semi-arid environment of the Mediterranean basin, where they are considered a traditional crop. Cardoons are in fact grown as vegetables in southern Europe and north Africa, and consumed for their whitened fleshy stalks; flowers are harvested and used as vegetable rennet
[4].
Over the last 30 years, the potential of cultivated cardoon for biomass production has been increasingly investigated
[5][6]. Advantages are represented by its perennial nature, the ability to photosynthesize during winter, and the high yields of lignocellulosic biomass (approximately 5–30 t/ha/year of dry matter, reviewed by
[7]) obtained with minimal agricultural inputs. Moreover, cardoon is a highly adaptive species able to tolerate abiotic stresses such as soil salinity
[8][9], drought
[10], and soil contamination by heavy metals
[11][12].
The use of cardoon to produce energy has been thoroughly evaluated
[13][14][15][16]; a recent study reported that the total energy (E) obtainable from cardoon biomass was 236 GJ/ha/year, with an increase of E measured over the three cropping years of the experiment, suggesting that the use of this species as an energy-crop might be effective especially in semi-arid areas with low productivity soils
[17]. Cardoon stalks have also been considered as a good source of fiber for the production of paper
[18].
Cardoon biomass should also be considered for the high content in specialized metabolites (SMs); these molecules are in fact high-value compounds in a biorefinery perspective. Notably, cardoon extracts, especially from leaves, are rich in phenylpropanoids
[19]. The most representative extracts are hydroxycinnamic and caffeoylquinic acids (particularly chlorogenic acid, CGA),
p-coumaric acid, and several flavonoids (vitexin, luteolin, apigenin, naringin)
[20][21][22]; these molecules mainly contribute to the antioxidant properties of the cardoon extracts
[23][24].
For example, CGA has been described as a bioactive molecule against obesity, diabetes, cancer, and for its role in cardio and neuro-protection
[25]. Moreover, cardoon extracts have potential in the prevention of hepatic and cardiac oxidative stresses
[26][27], and they have been studied for their anti-microbial and anti-fungal activities related to the presence of flavonoids
[24][28]. Therefore, the recovery of such nutraceuticals would be impactful for their possible downstream uses.
The importance of profiling phenylpropanoids is noteworthy when it comes to lignin. The term lignin refers to a large class of structural phenolic polymers that create complexes with the polysaccharides of cell walls (cellulose, hemicellulose and pectins); the abundance and chemical nature of such complexes contribute to the recalcitrance to the microbial or enzymatic degradative process of lignocellulosic biomass
[29]. Therefore, chemical/physical or biological pre-treatments are required to improve the digestibility and availability of the cellulose fraction to enzymatic degradation, and facilitate the recovery of fermentable sugars (saccharification). However, such pre-treatments are expensive and invasive, and they determine a reduction in the sustainability of biomass valorization
[30]. Consequently, using plant biomass with reduced and/or easily removable lignin is advantageous, and efforts have been performed to reach this goal through biotechnological approaches
[31].
Another interesting feature of cardoon biomass is represented by its oil content (largely derived from seeds); cardoon oils, given their high unsaturated to saturated fatty acids ratio, the high levels of oleic and linoleic acids, and stability to oxidation
[22][32], have potential for nutraceutical use, lowering serum cholesterol levels
[33], and for biorefinery, e.g. for the production of bioplastics
[34]. Indeed, plant oils and fats are among the most important renewable raw materials of the chemical industry, and examples of industrial applications include their use in environmentally friendly fluids and lubricants
[35]. More recently, the possibility of functionalizing fatty acids skeletons through organic synthesis and/or microbial reactions gave strong emphasis to the possibility of improving oil production in plants
[36]. Consequently, modification of the fatty acid flux, especially in favor of high monounsaturated fatty acids (MUFAs) accumulation, represents a major goal for biotechnology oriented towards green chemistry.
Among the drawbacks of cardoon biomass production from field cultivation, there are the variability in yields
[37] and in the quality of the lignocellulosic biomass
[38]. Field production also affects SMs: most of the cardoon genotypes assessed suffer from strong environmental effects on their phenolic profiles
[39], and, in particular, alterations of the phenylpropanoid levels can be traced back to biotic and abiotic stressors
[40]. Similarly, differences in the fatty acid profile of oils have been described as a result of genotype, plant tissues, climatic factors, and geographical regions
[22][32][41][42]. For these reasons, the development of cardoon cell cultures for biorefinery applications is a promising strategy alternative to field production. Undifferentiated plant cell cultures (PCC) are totipotent, i.e., able to express the full genetic machinery of their species/tissue of origin. Characterization and improvement of PCC is thus an important target of research, to overcome limitations imposed by climatic changes and/or pathogen attacks, and by the genetic segregation of traits of interest that are cancelled with the propagation of selected stable cell lines of clones
[43][44]. Moreover, many nutraceuticals are usually accumulated in low concentration in plant organs; the use of PCC and bioreactors, coupled with the possibility of selecting the best lines (and modifying their genome) and with elicitation techniques, improves the economic gains derived from the production, while at the same time reducing the environmental impacts bound to field cultivation
[45][46]. Furthermore, PCC are devoid from the risk of containing toxic substances, such as residual pesticides. Altogether, this paves the way to the development of a pipeline of great interest to use PCC in the frame of biorefinery
[44][47].
2. Development of a Method for the Stable Transformation of Cardoon
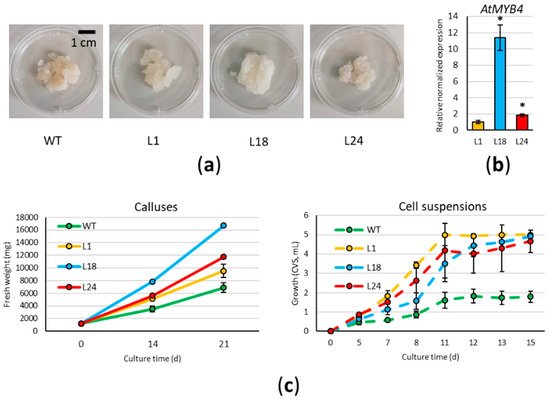
In this work, we developed a method for the stable genetic transformation of cardoon leaf-derived cell cultures via A. tumefaciens, overexpressing the coding sequence of the A. thaliana MYB4 transcription factor under the control of the constitutive cauliflower mosaic virus (CaMV35S) promoter. Five distinct transformation experiments were carried out: wild type (WT) calluses obtained from cardoon plants (“Spagnolo” genotype) were co-cultivated with AGL1 or EHA105 A. tumefaciens strains carrying the p35S::AtMYB4 construct. To select transformants, the co-cultivated cardoon cells were spread on the selective solid Gamborg B5 medium containing 10 mg/L phosphinothricin (PPT) and 200 mg/L cefotaxime (CFX), sub-culturing emerging calluses to new medium every 21 days. While at first, we counted 48 independent calluses emerging from the selective plates, only 15 of them survived the second and further sub-culturing rounds. Three months after transformation, all 15 resistant calluses were checked via PCR, and confirmed to be carrying the transgene and devoid of Agrobacterium contamination. All lines were maintained on PPT and CFX-containing media in order to avoid genetic chimerism, and the three AtMYB4oe lines selected to be analyzed for this study are here referred as L1, L24 (from infection with the AGL1 strain), and L18 (from infection with the EHA105 strain). The three lines appeared similar to each other, but different from the WT, with the callus texture softer than WT and thus more easily spreadable when sub-cultured (Figure 1a). RT-qPCR was used to verify the expression of AtMYB4, showing that the different lines expressed the transgene at different levels, with L1 having the lowest and L18 the highest level of expression (Figure 1b).
Figure 1. Phenotype and growth of cardoon lines: (a) Calluses of wild type (WT) and AtMYBoe lines (L1, L18, L24) grown for 21 days on solid medium; (b) Relative expression level (mean ± standard deviation, S.D.) of the transgene in the three lines selected for this study. Asterisks indicate significant differences (p < 0.01); (c) Growth curves for calluses grown on solid medium (left) and for cell suspension grown in liquid medium (right). CVS: cell volume after sedimentation. Each value represents the mean of three biological replicates ± S.D.
3. AtMYB4oe Lines Have Higher Growth Rates Than Wild Type
Growth rates of WT and AtMYB4oe lines were monitored both on solid and liquid media (Figure 1c); for this purpose, the starting material belonged to the XXII round of sub-cultures for WT, and to the XIV for AtMYB4oe lines. Calluses (6 × 200 mg each) were placed on Gamborg B5 solid medium in Petri dishes, and the total cellular mass (fresh weight, FW) was evaluated at 14 and 21 days of the sub-culture. All transgenic lines showed a significant higher growth rate than WT at both time points; in particular, after 14 days, AtMYB4oe biomass was 1.5-fold (L1 and L24) and 2-fold (L18) higher than WT. A similar trend was observed after 21 days in L1 and L24, whereas L18 mass increased 2.4-fold with respect to the WT. The fastest growing lines (L18 and L24) were also characterized by the lowest variation among experimental replicates, granting a more uniform growing behavior compared to WT. Relative differences of FW observed between WT and AtMYB4oe lines were confirmed in the comparison of dry weights (DW), as the percentage of the ratio between DW and FW was nearly identical for all samples (3.6%). Cell suspensions were grown for a maximum of 15 days, and the cell volume after sedimentation (CVS) was used to follow their growth. During the initial lag phase up to the fifth day, we observed a similar growth in all lines, whereas during the exponential phase, from the 5th to the 11th day, all the transgenic lines had a higher growth rate than the WT one (biomass 2–3-fold higher). L18 and L24 showed a slightly lower growth rate than L1 at the end of the exponential phase; nevertheless, on the 15th day, all the transgenic lines reached a biomass 3-fold higher than the WT.
Additionally, as plants of the “Spagnolo” genotype are tolerant to salinity stress
[9], we grew WT and transgenic lines, adding 100 mM NaCl to solid medium. WT line showed some degree of tolerance to this condition, whereas
AtMYB4oe lines did not grow, and turned quickly to necrosis.
4. AtMYB4oe Lines Show Decreased Phenolic Compounds and Lignin Content and Enhanced Enzymatic Saccharification Efficiency
To investigate if the overexpression of
AtMYB4 exerted a similar repression on the phenylpropanoid pathway to what was observed in other species, particularly on lignin, and as recently demonstrated also on anthocyanins and proanthocyanidins
[48], we measured the amount of phenolic compounds in extracts of 21 days-old freeze-dried calluses (sub-cultures: XXII for WT and XIV for
AtMYB4oe lines).
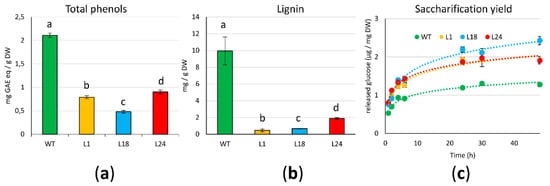
Figure 2a shows that all
AtMYB4oe lines have significantly less phenolic compounds than WT, with the strongest reduction in L18. As lignin monomers are one of the main products of the phenylpropanoid pathway, we quantified lignin content. As for total phenolic compounds, we reported a significant lignin reduction (close to −90%) for all transgenic lines (
Figure 2b).
Figure 2. Phenols/lignin content and saccharification yields of 21 days-old calluses: (a) Folin–Ciocalteu’s quantification of total phenols, expressed as mg of GAE (Gallic Acid Equivalents)/g dry weight (DW) ± S.D; (b) quantification of lignin via thioglycolic acid (TGA) derivatization. Bars indicate means ± S.D. of three biological replicates. Different letters above bars indicate statistically significant differences (p < 0.01); (c) saccharification yields expressed as mean values of µg released glucose/mg DW ± S.D.
Next, we wanted to test if the depletion in lignin content of AtMYB4oe lines had an impact on the saccharification yields of the lignocellulosic biomass, measured as the amount of fermentable sugars obtained from the hydrolysis of the cell wall polysaccharides. This provides an estimate of the accessibility of the cell wall to degradative enzymes and, consequently, of the accessibility of biomass to extract useful compounds. To do this, we measured over time the amount of glucose released from the freeze-dried biomass subjected to enzymatic degradation with a cellulase (Figure 2c). Over 48 h of reactions, logarithmic regression models fit very well with saccharification trends for WT and transgenic lines (R2 > 0.97); the amount of released glucose from AtMYB4oe lines was higher than WT already at the earliest time points of the reactions (1 h, 2 h), and this was also confirmed when the reaction plateau was reached. In fact, from 24 h on, L1 and L24 had a 1.3-fold increase of released glucose over WT, a value that is even higher (2-fold) in L18.
5. Transcriptomic Analysis
Transcriptomic changes induced by the heterologous overexpression of AtMYB4 were evaluated by RNA-seq analysis. A Gene Ontology (GO), followed by a Gene Ontology Enrichment Analysis (GOEA), was performed with the core-DEGs (defined as genes with log2 FC vs. WT > +/- 1 for all three lines) to identify the most enriched GO terms. The complexity of the list of enriched GO terms for biological processes (BP), molecular functions (MF), and cellular components (CC) was reduced via the software REVIGO, in order to identify and collapse redundant terms. Interestingly downregulated core-DEGs were categorized in processes such as lignin biosynthesis, phenylpropanoid metabolism, aromatic amino acids biosynthesis, hyperosmotic salinity response, and cell cycle arrest. On the other hand, BP relative to upregulated core-DEGs included the mitotic cell cycle, positive regulation of cell division, mitotic chromosome condensation, and microtubule-based movement.
6. Biochemical Profiles
WT and transgenic lines were analyzed for the characterization of phenolic compounds, antioxidant activities, and fatty acids (Table 1). This deep quantitative and qualitative characterization revealed interesting findings about specific nutraceuticals: CGA accumulates more than WT. Similarly, p-coumaric acid and flavonoids identified in the extracts (quercetin, kaempferol, naringin, luteolin, myricetin, and apigenin) accumulated at similar levels than WT. Notably, this confirms that while the overexpression of AtMYB4 represses the phenylpropanoid pathway by reducing the production of lignin monomers, it does not necessarily depauperate nutraceuticals, which could be recovered from the cell cultures, increasing the interest in these lines from a biotechnological point of view. Interestingly, we detected a 3–4-fold increase in L1 and L24 lines for oleic acid, whose increase in leves represents a goal of biorefinery approaches in many plant species.
Table 1. Biochemical characterization of WT and AtMYB4oe lines. Polyphenols content detected by HRMS-Orbitrap: values are expressed in ppm = µg/g dry weight, DW. 5-iFQA = 5-isoferuloyl quinic acid; 1,5-DiCQA = 1,5-dicaffeoyl quinic acid (cynarin); 3,4-DiCQA = 3,4-dicaffeoyl quinic acid; 5-FQA = 5-feruloyl quinic acid; 3-FQA = 3-feruloyl quinic acid; 3-CQA = 3-caffeoyl quinic acid (chlorogenic acid, CGA). Antioxidant activities were measured via 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,2’-azino-bis 3-ethylbenzothiazoline-6-sulfonic acid (ABTS) and Ferric Reducing Antioxidant Power (FRAP) assays. Values are expressed as TEAC: Trolox®-equivalent antioxidant capacity mmol Trolox/kg DW. Each value shown represents the mean values ± S.D. of three biological and two technical replicates. Fatty acids content was detected by GC-FID. Values are expressed as % of total; SFA = saturated fatty acids; MUFA = monounsaturated fatty acids; PUFA = polyunsaturated fatty acids. Different letters denote a significant difference among lines through analysis of variance (ANOVA). Statistical significance was defined as p < 0.05, using the Tukey’s post hoc test for mean separation.
| |
|
WT
|
L1
|
L18
|
L24
|
Polyphenols (µg/g DW)
|
5-iFQA |
21.50 ± 1.33a |
15.41 ± 0.43b |
0.11 ± 0.04c |
4.26 ± 0.12d |
| 1,5-DiCQA (cynarin) |
3345.79 ± 112.23a |
532.70 ± 11.55b |
2.50 ± 0.81c |
352.98 ± 21.71d |
| 3,4-DiCQA |
3152.41 ± 26.67a |
443.88 ± 12.43b |
103.58 ± 8.23c |
1615.65 ± 34.56d |
| 5-FQA |
98.87 ± 11.34a |
5.56 ± 0.43b |
0.83 ± 0.011c |
76.44 ± 11.34d |
| 3-FQA |
3.09 ± 0.55a |
2.71 ± 0.91b |
0.10 ± 0.01c |
10.20 ± 0.23d |
| 3-CQA (CGA) |
319.38 ± 22.45a |
424.66 ± 11.32b |
97.28 ± 3.56c |
513.48 ± 13.57d |
| p-coumaric acid |
3.50 ± 0.02a |
3.75 ± 0.91a |
1.50 ± 0.01b |
3.85 ± 0.02a |
| Quercetin |
3.35 ± 0.02a |
4.30 ± 0.02b |
0.55 ± 0.01c |
4.35 ± 0.02b |
| Quercetin-glucoside |
4.00 ± 0.65a |
5.25 ± 1.23b |
1.25 ± 0.03c |
5.40 ± 0.34b |
| Kaempferol |
3.55 ± 0.65a |
4.30 ± 0.91b |
1.05 ± 0.03c |
4.55 ± 0.83d |
| Kaempferol-3-O-glucoside |
1.15 ± 0.02a |
1.50 ± 0.03b |
0.40 ± 0.04c |
1.60 ± 0.04b |
| Naringin |
1.20 ± 0.01a |
1.75 ± 0.01b |
0.41 ± 0.03c |
1.800.03b |
| Luteolin |
1.50 ± 0.03a |
2.35 ± 0.02b |
0.60 ± 0.04c |
2.55 ± 0.04d |
| Myricetin |
2.55 ± 0.34a |
2.96 ± 0.32b |
0.73 ± 0.11c |
3.04 ± 0.12b |
| Apigenin |
0.20 ± 0.03a |
0.25 ± 0.02b |
0.01 ± 0.003c |
0.30 ± 0.01d |
| Total polyphenols |
6962.05a |
1451.33b |
210.92c |
2600.46d |
Antiox. activity (TEAC)
|
DPPH |
83.7 ± 0.61a |
22.17 ± 0.42b |
16.96 ± 0.63c |
35.43 ± 5.82d |
| ABTS |
71.33 ± 0.20a |
19.16 ± 0.07b |
12.54 ± 0.26c |
23.61 ± 0.22d |
| FRAP |
66.81 ± 0.98a |
30.84 ± 0.21b |
11.09 ± 0.08c |
27.90 ± 0.54d |
Oil %
|
|
11.38 |
8.24 |
10.35 |
7.25 |
Fatty acids (%)
|
Palmitic (C16:0) |
22.29 ± 1.12ab |
21.71 ± 0.58a |
24.06 ± 0.35b |
19.46 ± 0.38c |
| Stearic (C18:0) |
2.85 ± 0.31ab |
3.14 ± 0.54ab |
2.29 ± 0.07a |
3.73 ± 0.26b |
| Oleic (C18:1) |
3.43 ± 1.91a |
10.30 ± 0.56b |
3.49 ± 0.27a |
12.35 ± 0.16c |
| Linoleic (C18:2) |
19.38 ± 3.74a |
45.82 ± 0.85b |
41.04 ± 0.61c |
41.34 ± 0.39c |
| Linolenic (C18:3) |
42.60 ± 6.34a |
10.16 ± 0.38b |
20.73 ± 0.74c |
14.46 ± 0.06d |
| Arachidic (C20:0) |
0.80 ± 0.08a |
0.88 ± 0.01a |
0.84 ± 0.05a |
1.04 ± 0.05b |
| Lignoceric (C24:0) |
2.42 ± 0.30a |
3.26 ± 1.59a |
2.41 ± 0.58a |
2.40 ± 0.68a |
| Nervonic (C24:1) |
1.23 ± 0.14a |
0.18 ± 0.03b |
0.52 ± 0.47ab |
0.66 ± 0.12ab |
| Total SFA% |
30.54 ± 0.78ab |
30.29 ± 0.76a |
32.07 ± 0.43b |
28.00 ± 0.10c |
| Total MUFA % |
4.65 ± 1.777a |
10.48 ± 0.30b |
4.02 ± 0.21a |
13.01 ± 0.28c |
| Total PUFA % |
61.98 ± 2.88a |
55.98 ± 0.80b |
61.77 ± 0.41a |
55.81 ± 0.42b |
| Others % |
2.83 |
3.25 |
2.14 |
3.18 |
7. Conclusions and Future Perspectives
Authors have shown that the protocol we set up allows the stable genetic transformation of cardoon cells. The AtMYB4oe lines presented in this study proved to be valuable tools for their use in bioreactors. The main advantages of these lines are represented by their faster growth rate and improved accessibility of the biomass to enzymatic degradation, due to the reduction in lignin content, which also implies an easier extractability of compounds of interest, as well as an interesting modification in their nutraceutical value. The development of this technique represents a significant step towards the industrial use of cardoon cell cultures, which can be further improved targeting specific metabolic pathways of interest; two examples would be represented by targeting other MYB transcription factors involved in the production of specialized metabolites, or by the alteration of the activity of biosynthetic genes for fatty acids, both in the frame of gain or loss-of-function genetic approaches. Moreover, further exploration of the generated RNA-seq data could provide useful to further support molecular analyses of primary and specialized metabolic pathways of cardoon cell cultures. Finally, in order to evaluate whether the use of cardoon cells for biorefinery is energetically and economically sustainable, further studies on large-scale production are being conducted.
 +1 point
+1 point