Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Thierry Emonet | + 4316 word(s) | 4316 | 2021-07-08 04:33:05 | | | |
| 2 | Bruce Ren | -21 word(s) | 4295 | 2021-07-15 04:15:51 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Emonet, T. Non-Genetic Diversity in Chemosensing/Chemotactic Behavior. Encyclopedia. Available online: https://encyclopedia.pub/entry/12085 (accessed on 08 February 2026).
Emonet T. Non-Genetic Diversity in Chemosensing/Chemotactic Behavior. Encyclopedia. Available at: https://encyclopedia.pub/entry/12085. Accessed February 08, 2026.
Emonet, Thierry. "Non-Genetic Diversity in Chemosensing/Chemotactic Behavior" Encyclopedia, https://encyclopedia.pub/entry/12085 (accessed February 08, 2026).
Emonet, T. (2021, July 14). Non-Genetic Diversity in Chemosensing/Chemotactic Behavior. In Encyclopedia. https://encyclopedia.pub/entry/12085
Emonet, Thierry. "Non-Genetic Diversity in Chemosensing/Chemotactic Behavior." Encyclopedia. Web. 14 July, 2021.
Copy Citation
Non-genetic phenotypic diversity plays a significant role in the chemotactic behavior of bacteria, influencing how populations sense and respond to chemical stimuli.
bacterial chemotaxis
chemoreceptors
receptor clusters
phenotypic diversity
stochastic gene expression
fluctuations
signal processing
single cell
1. Introduction
Bacteria display very diverse morphologies and behaviors. This diversity allows genetically distinct individuals to serve different roles in a community. Within species also, genetic diversity can be an effective strategy to better utilize the environment [1], or evade antibiotic treatment, as is the case with some cancers and bacterial infections [2]. Interestingly, however, substantial phenotypic diversity occurs within populations of bacteria that share the same genome [3][4][5][6]. What is the molecular origin and functional consequences of such non-genetic phenotypic diversity?
Bacterial chemotaxis provides an effective quantitative framework to study non-genetic phenotypic diversity. In their seminal 1976 paper, Spudich and Koshland used the innovative “temporal gradient apparatus” [7], which combined a rapid mixing device with stroboscopic imaging, to characterize the swimming behavior of individual Salmonella bacteria in response to pulses of attractant [8]. Surprisingly, behavioral differences between individuals were greater than could be explained by potential mutations accumulating during growth. This and other early work in Escherichia coli [9][10] raised questions about cell-to-cell variability in chemotactic behavior and how collective migration occurs in the midst of phenotypic diversity. Since then, non-genetic diversity has been studied extensively in many biological systems, revealing fundamental mechanisms that generate it, including stochastic gene expression and uneven partitioning of biomolecules at cell division [11][12][13][14]. For more general information on the molecular mechanisms and functional consequences of cell-to-cell variability, we refer the reader to these reviews [3][4][13][15][16].
Here, we focus on non-genetic diversity in chemical sensing and the well-characterized chemotaxis system of E. coli for which there is a relatively good understanding of the molecular mechanism by which the chemotactic signaling pathway uses external signals to bias cell’s run-and-tumble motility toward favorable locations [17][18]. In this two-component system, five types of chemoreceptors (Tar, Tsr, Trg, Tap, and Aer) form homodimers arranged in trimers of dimers, themselves arranged in hexagonal latices comprising thousands of receptors connected by the scaffold protein CheW and the histidine kinase CheA [19][20][21]. The receptor-kinase complexes are highly cooperative at the level of the CheA activity [22], enabling signal amplification. Upon binding an attractant, receptor-kinase complexes switch to an inactive conformation, reducing the rate of phosphorylation of the response regulator CheY. Due to the phosphatase activity of CheZ, phosphorylated CheY-p concentration rapidly (<1 s) decreases [18], leading to a reduction in the probability to tumble. The prestimulus level of tumbling frequency is later reestablished via the action of CheR and CheB, which methylates and demethylates the cytoplasmic tail of the receptors when they are in the inactive and active conformations, respectively.
The general features of chemotaxis [17][18][20][23] and its quantitative treatment [18][24][25] have been reviewed elsewhere recently. Here, our intent is to review the current understanding of non-genetic phenotypic diversity in E. coli chemotaxis and how knowledge of phenotypic diversity affects our general understanding of chemosensing and chemotactic behavior.
2. Molecular Mechanisms Underlying Phenotypic Diversity in E. coli Chemotaxis
Quantifying phenotypic diversity first requires quantifying phenotypes, which can be accomplished by measuring functional and behavioral parameters in individual cells. For E. coli chemotaxis, such parameters include the following: the tumble bias—the fraction of time the cell spends tumbling; the switching frequency—the number of switches per unit of time between the run and tumble states; the pathway gain, or magnitude of the response relative to a stimulus change both at the level of the kinase and of the motor; the adaptation time, or time to return to the original behavioral pattern after a stimulus; and the rotational diffusion, all of which affect chemotactic performance [10][18][25][26][27][28][29][30]. Alternatively, the phenotype of an individual cell can also be quantified without making assumptions about the set of possible behavior states a bacterium can be in (e.g., run-and-tumble). In one approach, each individual cell trajectory was represented as a scatter plot of time points in the space of translational and rotational velocity [31]. Viewed as a two-dimensional probability distribution, the scatter plot defines the motility states the bacterium exhibit over time and the fraction of time it spends in each [32][33].
An important consideration is that phenotypic parameters may vary on multiple time-scales. For example, a cell might exhibit the same tumble bias over its entire life span, or its tumble bias might change from the moment it is born until it divides. At any given instant, multiple stochastic processes operate on different time scales and contribute to phenotypic variability (Figure 1). Below, we review recent advances in current understanding of these various mechanisms and how they affect E. coli chemotaxis.
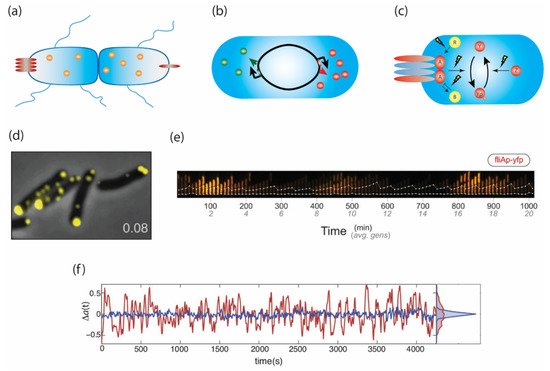
Figure 1. Phenotypic diversity arises from processes operating at different timescales. (a) Unequal partitioning of proteins and organelles upon cell division, (b) stochastic gene expression, and (c) stochasticity in the chemical reactions of signaling pathways all contribute to phenotypic diversity. (d) The number and size of chemoreceptor clusters varies from cell to cell due to random partitioning. This variation may lead to diverse sensitivities to stimuli. (e) The expression of class II motility genes such as fliA occurs in pulses lasting multiple generations. These pulses are stochastic, likely generating cells with very different sensory and swimming capabilities. (f) Fluctuations in the kinase activity of the chemotaxis network. In red are cells containing the adaptation enzymes CheR and CheB, while in blue are cheRB deletants. The cycle of methylation and demethylation, as well as receptor cooperativity both contribute to fluctuations. Panels d, e, and f are adapted from Koler et al., 2018 [34]; Kim et al., 2020 [35]; and Keegstra et al., 2017 [36].
2.1. Variation Arising at Cell Division
One mechanism likely to contribute to long-lasting differences between cells is partitioning noise—the unequal distribution of biomolecules between daughter cells during cell division (Figure 1a). General features of partitioning noise and its quantitative treatment have been subject to review elsewhere [13][37]. For chemosensing and motility, the partitioning of organelles, such as molecular motors and large receptor clusters, is likely to contribute to phenotypic variability. However, so far, only few studies have examined this aspect of phenotypic variability.
Recent studies have determined the location and size distributions of chemoreceptor clusters in growing cells [38][39]. At early growth stages, cells contain multiple small receptor clusters, which are positioned along the length of the cell (Figure 1d) [34]. However, as the culture approaches stationary phase, individual cells typically contain only one or two polar clusters. The resulting cell-to-cell differences in cluster size and number are likely to translate into differences in sensitivity to chemical signals. While a direct link between cluster positioning and portioning noise has not been demonstrated directly, an uneven distribution of chemoreceptor complexes could contribute to long-lasting diversity, since some individuals will inherit multiple clusters, while others will inherit none.
2.2. Stochastic Pulses of Motility Gene Expression
By far, the most-studied source of phenotypic variability is stochastic gene expression (Figure 1b), where it is understood that transcription and translation events occur in bursts with variable frequency and size, leading to broad distributions of protein expression in isogenic cell populations [6][11][12][14]. Different regulatory schemes can lead to fluctuations in gene expression with various magnitudes and timescales [40][41]. In E. coli, the arrangement of chemotaxis genes on multicistronic operons has been shown to buffer the effect of some of these fluctuations in the signaling output by ensuring gene products that participate in the same step of signal transduction are kept at an acceptable ratio [42]. Furthermore, a temperature-sensitive secondary RNA structure upstream of cheR was suggested to compensate for temperature-induced differences in the rate of adaptation of cells to signals [43][44]. While these compensatory mechanisms ensure that individual phenotypes remain functional, gene expression noise gives rise to large cell-to-cell variability within this range of functional phenotypic space [45]. For example, the expression ratio of different receptor species varies greatly [46], leading to variation in the sensitivity for different chemoeffectors [36][47].
Recently, a study by the Cluzel lab revealed how regulated stochasticity in gene expression plays an important role in the transcription network governing E. coli chemotaxis [35]. In E. coli, there are 14 operons collectively encoding the flagellar components and the chemotaxis machinery. These operons contain over 50 genes that are regulated in a three-tier hierarchy. At the top is the class I operon, flhDC, which encodes the master regulator of flagella and chemotaxis. Then, FlhDC activates the class II genes encoding the basal body and flagellar hook, as well as the alternative sigma factor FliA, which subsequently activates the class III genes encoding the flagellar filament and chemotaxis network [48].
Using a microfluidic device called the “mother machine” where a single cell’s lineage can be monitored over multiple division events, the Cluzel lab quantified the expression of class I, II, and III genes over multiple generations using fluorescent reporters [35]. While the class I genes were expressed constitutively, class II and III gene expression occurred in pulses lasting multiple generations interspersed with long periods of inactivity (Figure 1e). Interestingly, these pulses did not depend on the regulation of flhDC transcription but were instead shown to require interactions between FlhDC and its post-translational regulator YdiV, which amplified transcriptional noise, allowing small temporal fluctuations in flhDC transcription to generate large pulses of class II expression. In short, transcriptional noise, coupled with a post-translational circuit, converts constitutive expression of class I genes into pulsatile expression of class II and III genes.
The pulsatile expression pattern of class II genes was further explored in theoretical work that proposed a possible molecular mechanism [49]. Simple stochastic models of class I and class II gene expression successfully capture the expected distributions of class II expression. By fitting these models to single-cell data [35] of class II expression with and without YdiV, Sassi et al. found that YdiV’s functions were twofold: first, it makes class II gene expression ultrasensitive to changes in FlhDC concentration. Second, it allows the system to integrate FlhDC expression over time to filter away small fluctuations. As such, they proposed a molecular mechanism where YdiV sequesters FlhDC, such that small changes in FlhDC expression lead to ultrasensitive increases in class II promoter activity [49]. This simple sequestration mechanism allows the expression of motility genes to follow a filter-and-integrate method of pulse generation that is qualitatively distinct from other known biological pulse generators that rely on bistability in transcriptional feedback loops [50][51].
What is the end result of such an expression pattern? Ultimately, stochastic gene expression generates a wide diversity of sensory and motile phenotypes that may contribute to phenotypic changes over the course of cell growth as observed in behavioral studies [33]. An interesting hypothesis is that cells in generations following a valley of little expression could have different chemotactic phenotypes than those following a pulse of expression. Such an expression pattern could also play a role in how individual cells within one population sort themselves in space during collective migration, as described in later sections.
2.3. Spontaneous Temporal Fluctuations in Pathway Activity
In addition to partitioning and gene expression noise, the behavior of individual cells is also affected by temporal fluctuations that arise due to the inherent stochasticity of the chemical reactions of the chemotaxis signal transduction pathway (Figure 1c). The first study to characterize these fluctuations measured the rotational direction of latex beads attached to flagella in cells that were adapted to a motility buffer devoid of any signal [52]. Analyzing the power spectra of these time-series revealed surprisingly large fluctuations in tumble bias with characteristic times scales on the order of 10s of seconds. Blocking adaptation by receptor methylation eliminated these fluctuations, while titrating the amount of methyltransferase CheR changed their time scale and amplitude [52].
These data can be recapitulated with a coarse-grained model of the signaling pathway in which the rates of methylations and demethylations follow Michaelis–Menten kinetics with a sub-linear dependency on the activity of the receptors [45][52][53][54][55]. This model assumes that in vivo, CheR and CheB work near saturation [53][54], which is consistent with population average FRET measurements of adaption kinetics [56] and with stoichiometric measurements—CheRB are expressed orders of magnitude less than the receptors [57]. A consequence of the saturated enzyme kinetics hypothesis is that the steady-state activity of the system becomes ultra-sensitive [58] to the ratio of CheR and CheB abundances, which can contribute to the amplification of spontaneous fluctuations in the activity of the receptors [45][52][54][55].
Recently, the wide availability of high-sensitivity cameras allowed the classic CheZ/CheY FRET method [59]—originally developed to measure kinase activity in a population of cells—to be adapted for quantifying kinase activity in single cells [36][60][61][62] (Figure 1f). Measurements of the kinase activity in individual unstimulated cells revealed large fluctuations in the kinase activity that could be attributed to two different sources. The first source was traced back to the activity of CheR and CheB. Similar to previous results [52], these fluctuations have characteristic time scales of tens of seconds and were strongly suppressed in cheRB-. Moreover, perturbations to the CheB phosphorylation feedback loop caused changes in steady-state kinase activity that were consistent with an ultrasensitive dependency of the activity of receptor-kinase complexes on the ratio of CheR/CheB abundances, and the role of the CheB phosphorylation feedback loop in reducing such ultra-sensitivity [36][54].
Interestingly, the magnitude of the fluctuations measured at the kinase were larger than previous estimates from fluctuations measured at the motor output [36], suggesting that a second source of fluctuations might be present. Indeed, measurements in cheRB- cells stimulated with a constant sub-saturating stimulus revealed methylation-independent fluctuations with time scales of 100s of seconds [36][60]. These fluctuations were eliminated when the scaffolding protein that mediates allosteric interactions in the receptor-kinase complexes, CheW, was replaced with the mutant CheW-X2 [60], which is a mutant that abolishes allosteric interactions but maintains signaling capabilities [22][63]. These large fluctuations also depended on the composition of receptor clusters and became almost switch-like between a few discrete states in cells that only expressed Tsr [36]. Together, these data suggest that the second source of fluctuations is caused by thermal fluctuations in highly cooperative receptor clusters [36][60].
While a full biophysical model is still missing, the emerging picture is that receptor clustering and adaptation kinetics both contribute to temporal fluctuations in the chemotaxis pathway. While thermal fluctuations and allosteric interactions in receptor clusters can introduce large fluctuations over very long timescales (up to 100s of seconds), the stochasticity of methylation events contributes smaller fluctuations on time scales of 10s of seconds. Interestingly, receptor clustering is required for fluctuations to be observed even in the presence of methylation and demethylation, which is probably because of the role of “assistance neighborhoods”—in which CheR (or CheB) tether to one receptor and methylate (or demethylate neighboring receptors—in signal amplification and adaptation kinetics; see [60][63][64][65][66]. The ultra-sensitivity of the kinase activity with respect to the ratio of CheR/CheB is also a source of phenotypic variability, which becomes more apparent when the buffering effect of the CheB phosphorylation feedback is removed [36].
3. Functional Consequences of Phenotypic Diversity on Chemosensing and Chemotactic Performance
Phenotypic diversity can be beneficial for populations of bacteria, especially in the face of environmental uncertainty [3][4][5][67]. In this section, we review how phenotypic diversity impacts chemotactic performance, how fluctuations in the chemotactic signaling pathway affect a cell’s information-processing ability, and how cell populations perform collective behaviors despite containing a multitude of sensory and behavioral phenotypes.
3.1. Functional Consequences of Phenotypic Diversity
Populations of E. coli display widely diverse swimming behaviors. Some of the first studies of run-and-tumble motion in E. coli demonstrated that adaptation time [8], tumble bias, and swimming speed [10] vary from cell to cell. New methodologies in single-cell tracking have expanded on these measurements, allowing precise, high-throughput quantification of single-cell behavioral parameters including the run-speed, turning angle, and tumble bias [26][27][29][30][31]. Meanwhile, optical tweezers have provided high time-resolution measurements of adaptation in single cells [68][69].
How do the processes described in previous sections contribute to this diversity? Variable expression of flagellar components, chemoreceptors, and sensory pathway components may all be relevant for certain aspects of E. coli’s behavioral diversity. For example, variation in the CheR/CheB ratio leads to a variation in tumble bias. Interestingly, the variance of this distribution depends primarily on CheB expression and not CheR, suggesting that mutations in regulatory elements might allow populations to adapt both the mean and variance of tumble biases by changing CheB and CheR expression [31][45]. Variations in tumble bias has profound consequences on chemotactic performance. By tracking individual cells in a static gradient of attractant, Waite et al. measured the relationship between tumble bias and drift velocity of individual cells [70]. They found that in liquid, wild-type cells with low tumble bias (TB = 0.01) climb gradients much faster than cells with intermediate tumble bias (TB = 0.2), as predicted by theory [71]. Next, by titrating the level of expression of CheR (or CheY) in the population, they could establish a causal relationship between CheR (or CheZ), tumble bias, and gradient climbing speed. Additionally, variation in pathway gain has been inferred from both single-cell tracking [28] and a microfluidic T-maze that sorts the population by chemotactic drift speed [72]. Diversity in pathway gain may result from diversity in the expression of different receptor species [46].
One way in which the diversity of sensory and swimming phenotypes benefits bacteria is to increase the overall chemotactic performance of the population beyond that of the mean phenotype within the population. This was apparent in Waite et al.’s experiment mentioned above. At approximately wild-type CheR expression, the motion of the population’s mean position closely followed the position of cells with average tumble bias. However, at lower average CheR expression, the population’s mean position ascended the gradient more quickly than cells with average tumble bias. This discrepancy between population performance and the performance of the mean phenotype can be explained by Jensen’s inequality [73]: for a convex function of a random variable f(x)
, the average of value of the function with respect to the random variable x, E[f(x)], is equal or greater than the value of the function evaluated at the average x, f(E[x])
. Since the relationship between phenotype and gradient-climbind speed is nonlinear at low CheR expression and linear at high CheR expression, populations in the low expression regime climb gradients more quickly on average when there is variation in tumble bias [70].
In addition to directly modulating population performance, phenotypic diversity can allow populations to survive in diverse, fluctuating environments [3][5]. Bet-hedging is a general ecological strategy where populations generate individuals with phenotypes poorly adapted for the current environment but well adapted for a different environment. This strategy ensures the population’s survival after an environmental change [74][75]. Theoretical work in E. coli chemotaxis demonstrated that variability in tumble bias and adaptation time allows the population to navigate toward nutrients appearing at different distances away from the population [45]. Since a single phenotype cannot optimally locate both near and far attractant sources [76], generating diverse phenotypes is necessary for the population’s survival in the wild, where nutrients may appear any distance from the population.
An attractive hypothesis is that variation in behavioral parameters also allows E. coli to perform well in both liquid and porous solid environments, where the optimal phenotypes are likely different. New methods developed in the Datta lab are revealing how E. coli navigate disordered porous environments. Three-dimensional porous environments were constructed using small, clear, hydrogel particles, allowing for imaging of single bacteria in environments of different average pore diameter [77][78]. In such environments, cells do not employ a run-and-tumble strategy, as in unconfined liquid environments. Instead, they hop between openings in the media after changes in flagellar bundling allow them to escape confined spaces [77]. Efficient navigation in such an environment could require a different tumble bias than in liquid media; as such, it is possible that phenotypic diversity allows the population to bet-hedge against the physical properties, as well as the chemical makeup, of the environment.
3.2. Consequences of Temporal Variation in the Chemosensory Pathway
As discussed above, behavioral variation within populations can be beneficial; are signaling fluctuations within individual cells similarly beneficial? Such fluctuations corrupt the transmitted signal [79]. Since the system consumes energy (in the form of ATP for CheY phosphorylation and S-adenosyl methionine for methylation), it should in principle be possible to decrease the magnitude of fluctuations, as shown in a recent theoretical analysis [80]. Why then has the system evolved to maintain these fluctuations?
One reason may be the multi-task nature of the chemotaxis signaling pathway: it must process external signals to bias chemotactic behavior in the presence of attractant gradients, but it must also drive swimming behavior in the absence of gradients to find better conditions for growth. Since the first observation of signaling fluctuations [52], it has been suggested that such fluctuations lead to fat-tailed distributions of run lengths over a few decades [54][81]. A random walk with such a distribution of run-lengths approximates what is called a ‘Levy flight/walk’. In a Levy walk, long runs occasionally bring the cell to new, unexplored territory, leading to more efficient exploration of the environment [82]. Numerical simulations have also shown that Levy-walk behavior helps cells respond to and climb shallow chemoattractant gradients more efficiently by increasing the coordination between individual flagellar motors and enabling cells to detect very small signals by integrating stimuli over long durations [83][84].
Despite intensive theoretical study of the causal relationship between signaling fluctuations and Levy walks of free swimming cells [81][82], experimental evidence that E. coli performs a Levy walk has remained circumstantial. Recently, however, three-dimensional tracking of individual bacteria over a long time have revealed how in the absence of chemoattractant, signaling fluctuations drive long tails in the distribution of run durations [26]. Comparing wild-type behavior with that of a mutant strain lacking signaling noise indicates that a consequence of the long tails in run duration is a superdiffusive swimming behavior consistent, at least over a decade and a half, with Levy walk [85] (Figure 2a). Levy flight has also been observed in swarming bacteria [86]. Thus, fluctuations in the chemotaxis pathway can modulate behavior for efficient spatial exploration over multiple length scales.
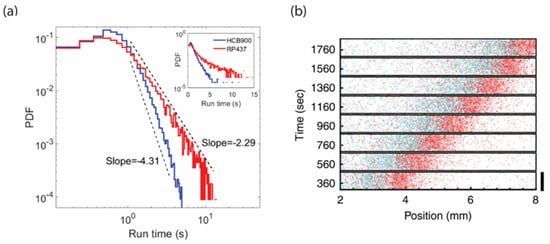
Figure 2. Phenotypic diversity and signaling fluctuations affect ecological performance. (a) The distribution of run lengths in wild-type cells (red) has a fatter tail than in cells with the same mean tumble bias but no fluctuations in CheY phosphorylation (blue). This distribution of run lengths allows cells to efficiently explore the environment by Levy walk instead of a pure random walk. (b) The conflict between individuality and collective behavior is resolved by spatial sorting of phenotypes in traveling waves of bacteria. Cells with lower tumble bias (red) climb attractant gradients more quickly than cells with higher tumble bias (blue), and therefore are located near the front of the wave. Panels a and b are adapted with permission from Huo et al., 2021 [85]; Fu et al., 2018 [87].
The multi-task nature of the chemotaxis signaling network raises the question of whether the system still effectively operates as an information-processing system in biasing run-and-tumble behavior. Combining theory and quantitative measurements, Mattingly and Kamino et al. measured information-theoretic properties of the E. coli chemotaxis pathway for cells climbing shallow gradients [62]. Due to the presence of the signaling fluctuations, the amount of information E. coli acquired from the environment during chemotaxis was quite low—on the order of 0.01 bits/s in a shallow gradient where chemoattractant concentration varies on centimeter scale. This is far less than the 1 bits/s the cell needs to determine within a 1 s run whether it is swimming up or down of the gradient. However, the same experiments revealed that E. coli used that little information efficiently, achieving drift speed up the gradient around 70% of the theoretical maximum achievable performance. Thus, in spite of the large signaling noise, the chemotaxis signaling pathway seems to have been selected for the efficient usage of information it gathers from the environment.
3.3. Spatial Sorting of Chemotaxis Phenotypes
Despite containing diverse sensory and swimming phenotypes, populations of E. coli can perform collective migration, moving as a cohesive wave that chases a front of attractant generated by the cells consuming the environment [9][88]. How can populations achieve collective behavior in the midst of phenotypic diversity?
Recent work that combined theory and experiments conducted in microfluidics revealed that population migrating in an Adler wave form resolves the conflict between individuality and collective behavior by spontaneously organizing the position of the different phenotypes within the traveling wave [87]. It turns out that not all positions in the wave are equally difficult to navigate. Near the front, the gradient is shallower, and therefore, chemotaxis is more difficult. However, further back, navigation is easier, since the gradient is steeper due to a higher density of cells consuming the environment at that position. These position-dependent differences in signal strength cause the more performant cells to locate at the front where the signal is weaker and the lower performers to locate near the back where the gradient is steeper. This spatial organization can be visualized using mixtures of cells that express CheZ at different levels: CheZ controls tumble bias, which in turn affects the speed at which a cell can climb a gradient [70]. The cells with the higher tumble bias (lower level of expression of CheZ) are located behind those with lower tumble bias (red and blue dots in Figure 2b). Moreover, the distance between the peaks of the two populations traveling together increased with the distance in phenotypic space. Thus, phenotypic diversity leads to spatial sorting withing migrating groups of bacteria.
At the very back of the wave, the gradient becomes shallow again, and the concentration of signal eventually goes below detection level. Thus, phenotypes located near the back are at a higher risk of falling behind the migrating front, leading to a slow leakage of cells of the weakest phenotype at the back of the wave. Recent studies of chemotactic populations find that expanding populations are characterized by a group of pioneer cells that explore new environments and leave behind settler offspring that grow [1][89]. That is to say, bacteria may generate both chemotactic and sessile individuals as part of a division of labor strategy [15][90][91][92] to best utilize their environments.
References
- Liu, W.; Cremer, J.; Li, D.; Hwa, T.; Liu, C. An evolutionarily stable strategy to colonize spatially extended habitats. Nature 2019, 575, 664–668.
- Balaban, N.Q.; Helaine, S.; Lewis, K.; Ackermann, M.; Aldridge, B.; Andersson, D.I.; Brynildsen, M.P.; Bumann, D.; Camilli, A.; Collins, J.J.; et al. Definitions and guidelines for research on antibiotic persistence. Nat. Rev. Genet. 2019, 17, 441–448.
- Ackermann, M. A functional perspective on phenotypic heterogeneity in microorganisms. Nat. Rev. Genet. 2015, 13, 497–508.
- Martins, B.M.; Locke, J.C. Microbial individuality: How single-cell heterogeneity enables population level strategies. Curr. Opin. Microbiol. 2015, 24, 104–112.
- Norman, T.M.; Lord, N.D.; Paulsson, J.; Losick, R. Stochastic Switching of Cell Fate in Microbes. Annu. Rev. Microbiol. 2015, 69, 381–403.
- Raj, A.; Van Oudenaarden, A. Nature, Nurture, or Chance: Stochastic Gene Expression and Its Consequences. Cell 2008, 135, 216–226.
- Macnab, R.M.; Koshland, D.E., Jr. The gradient-sensing mechanism in bacterial chemotaxis. Proc. Natl. Acad. Sci. USA 1972, 69, 2509–2512.
- Spudich, J.L.; Koshland, D.E. Non-genetic individuality: Chance in the single cell. Nature 1976, 262, 467–471.
- Adler, J. Chemotaxis in Bacteria. Science 1966, 153, 708–716.
- Berg, H.C.; Brown, D.A. Chemotaxis in Escherichia coli analysed by Three-dimensional Tracking. Nature 1972, 239, 500–504.
- Cai, L.; Friedman, N.; Xie, X.S. Stochastic protein expression in individual cells at the single molecule level. Nature 2006, 440, 358–362.
- Elowitz, M.B.; Levine, A.J.; Siggia, E.D.; Swain, P. Stochastic Gene Expression in a Single Cell. Science 2002, 297, 1183–1186.
- Huh, D.; Paulsson, J. Non-genetic heterogeneity from stochastic partitioning at cell division. Nat. Genet. 2010, 43, 95–100.
- Taniguchi, Y.; Choi, P.J.; Li, G.-W.; Chen, H.; Babu, M.; Hearn, J.; Emili, A.; Xie, X.S. Quantifying E. coli Proteome and Transcriptome with Single-Molecule Sensitivity in Single Cells. Science 2010, 329, 533–538.
- Giri, S.; Waschina, S.; Kaleta, C.; Kost, C. Defining Division of Labor in Microbial Communities. J. Mol. Biol. 2019, 431, 4712–4731.
- Takhaveev, V.; Heinemann, M. Metabolic heterogeneity in clonal microbial populations. Curr. Opin. Microbiol. 2018, 45, 30–38.
- Bi, S.; Sourjik, V. Stimulus sensing and signal processing in bacterial chemotaxis. Curr. Opin. Microbiol. 2018, 45, 22–29.
- Colin, R.; Sourjik, V. Emergent properties of bacterial chemotaxis pathway. Curr. Opin. Microbiol. 2017, 39, 24–33.
- Hazelbauer, G.L.; Falke, J.J.; Parkinson, J.S. Bacterial chemoreceptors: High-performance signaling in networked arrays. Trends Biochem. Sci. 2008, 33, 9–19.
- Parkinson, J.S.; Hazelbauer, G.L.; Falke, J.J. Signaling and sensory adaptation in Escherichia coli chemoreceptors: 2015 update. Trends Microbiol. 2015, 23, 257–266.
- Yang, W.; Briegel, A. Diversity of Bacterial Chemosensory Arrays. Trends Microbiol. 2020, 28, 68–80.
- Piñas, G.E.; Frank, V.; Vaknin, A.; Parkinson, J.S. The source of high signal cooperativity in bacterial chemosensory arrays. Proc. Natl. Acad. Sci. USA 2016, 113, 3335–3340.
- Karmakar, R. State of the art of bacterial chemotaxis. J. Basic Microbiol. 2021, 61, 366–379.
- Tu, Y. Quantitative modeling of bacterial chemotaxis: Signal amplification and accurate adaptation. Annu. Rev. Biophys. 2013, 42, 337–359.
- Waite, A.J.; Frankel, N.W.; Emonet, T. Behavioral Variability and Phenotypic Diversity in Bacterial Chemotaxis. Annu. Rev. Biophys. 2018, 47, 595–616.
- Figueroa-Morales, N.; Soto, R.; Junot, G.; Darnige, T.; Douarche, C.; Martinez, V.A.; Lindner, A.; Clément, É. 3D Spatial Exploration by E. coli Echoes Motor Temporal Variability. Phys. Rev. X 2020, 10, 21004.
- Grognot, M.; Taute, K.M. More than propellers: How flagella shape bacterial motility behaviors. Curr. Opin. Microbiol. 2021, 61, 73–81.
- Masson, J.B.; Voisinne, G.; Wong-Ng, J.; Celani, A.; Vergassola, M. Noninvasive inference of the molecular chemotactic response using bacterial trajectories. Proc. Natl. Acad. Sci. USA 2012, 109, 1802–1807.
- Taute, K.M.; Gude, S.; Tans, S.J.; Shimizu, T.S. High-throughput 3D tracking of bacteria on a standard phase contrast microscope. Nat. Commun. 2015, 6, 8776.
- Turner, L.; Ping, L.; Neubauer, M.; Berg, H.C. Visualizing Flagella while Tracking Bacteria. Biophys. J. 2016, 111, 630–639.
- Dufour, Y.S.; Gillet, S.; Frankel, N.W.; Weibel, D.B.; Emonet, T. Direct Correlation between Motile Behavior and Protein Abundance in Single Cells. PLoS Comput. Biol. 2016, 12, e1005041.
- Jordan, D.; Kuehn, S.; Katifori, E.; Leibler, S. Behavioral diversity in microbes and low-dimensional phenotypic spaces. Proc. Natl. Acad. Sci. USA 2013, 110, 14018–14023.
- Pleška, M.; Jordan, D.; Frentz, Z.; Xue, B.; Leibler, S. Nongenetic individuality, changeability, and inheritance in bacterial behavior. Proc. Natl. Acad. Sci. USA 2021, 118, 2023322118.
- Koler, M.; Peretz, E.; Aditya, C.; Shimizu, T.S.; Vaknin, A. Long-term positioning and polar preference of chemoreceptor clusters in E. coli. Nat. Commun. 2018, 9, 4444.
- Kim, J.M.; Garcia-Alcala, M.; Balleza, E.; Cluzel, P. Stochastic transcriptional pulses orchestrate flagellar biosynthesis in Escherichia coli. Sci. Adv. 2020, 6, eaax0947.
- Keegstra, J.M.; Kamino, K.; Anquez, F.; Lazova, M.D.; Emonet, T.; Shimizu, T.S. Phenotypic diversity and temporal variability in a bacterial signaling network revealed by single-cell FRET. eLife 2017, 6, e27455.
- Baptista, I.; Ribeiro, A.S. Stochastic models coupling gene expression and partitioning in cell division in Escherichia coli. Biosystem 2020, 193–194, 104154.
- Jones, C.W.; Armitage, J.P. Positioning of bacterial chemoreceptors. Trends Microbiol. 2015, 23, 247–256.
- Solari, J.; Anquez, F.; Scherer, K.M.; Shimizu, T.S. Bacterial Chemoreceptor Imaging at High Spatiotemporal Resolution Using Photoconvertible Fluorescent Proteins; Springer: New York, NY, USA, 2018; pp. 203–231.
- Brameyer, S.; Hoyer, E.; Bibinger, S.; Burdack, K.; Lassak, J.; Jung, K. Molecular Design of a Signaling System Influences Noise in Protein Abundance under Acid Stress in Different Gammaproteobacteria. J. Bacteriol. 2020.
- Jones, D.L.; Brewster, R.C.; Phillips, R. Promoter architecture dictates cell-to-cell variability in gene expression. Science 2014, 346, 1533–1536.
- Kollmann, M.; Løvdok, L.; Bartholomé, K.; Timmer, J.; Sourjik, V. Design principles of a bacterial signalling network. Nature 2005, 438, 504–507.
- Dufour, Y.S.; Sneddon, M.W.; Emonet, T. Thermal Robustness: Lessons from Bacterial Chemotaxis. Curr. Biol. 2011, 21, R465–R468.
- Oleksiuk, O.; Jakovljevic, V.; Vladimirov, N.; Carvalho, R.; Paster, E.; Ryu, W.S.; Meir, Y.; Wingreen, N.S.; Kollmann, M.; Sourjik, V. Thermal Robustness of Signaling in Bacterial Chemotaxis. Cell 2011, 145, 312–321.
- Frankel, N.W.; Pontius, W.; Dufour, Y.S.; Long, J.; Hernandez-Nunez, L.; Emonet, T. Adaptability of non-genetic diversity in bacterial chemotaxis. eLife 2014, 3, e3526.
- Yoney, A.; Salman, H. Precision and Variability in Bacterial Temperature Sensing. Biophys. J. 2015, 108, 2427–2436.
- Kalinin, Y.; Neumann, S.; Sourjik, V.; Wu, M. Responses of Escherichia coli bacteria to two opposing chemoattractant gradients depend on the chemoreceptor ratio. J. Bacteriol. 2010, 192, 1796–1800.
- Apel, D.; Surette, M.G. Bringing order to a complex molecular machine: The assembly of the bacterial flagella. Biochim. Biophys. Acta (BBA) Biomembr. 2008, 1778, 1851–1858.
- Sassi, A.S.; Garcia-Alcala, M.; Kim, M.J.; Cluzel, P.; Tu, Y. Filtering input fluctuations in intensity and in time underlies stochastic transcriptional pulses without feedback. Proc. Natl. Acad. Sci. USA 2020, 117, 26608–26615.
- Hao, N.; O’Shea, E.K. Signal-dependent dynamics of transcription factor translocation controls gene expression. Nat. Struct. Mol. Biol. 2011, 19, 31–39.
- Locke, J.C.W.; Young, J.W.; Fontes, M.; Jimenez, M.J.H.; Elowitz, M.B. Stochastic Pulse Regulation in Bacterial Stress Response. Science 2011, 334, 366–369.
- Korobkova, E.; Emonet, T.; Vilar, J.M.G.; Shimizu, T.S.; Cluzel, P. From molecular noise to behavioural variability in a single bacterium. Nature 2004, 428, 574–578.
- Barkai, N.; Leibler, S. Robustness in simple biochemical networks. Nature 1997, 387, 913–917.
- Emonet, T.; Cluzel, P. Relationship between cellular response and behavioral variability in bacterial chemotaxis. Proc. Natl. Acad. Sci. USA 2008, 105, 3304–3309.
- Park, H.; Pontius, W.; Guet, C.C.; Marko, J.F.; Emonet, T.; Cluzel, P. Interdependence of behavioural variability and response to small stimuli in bacteria. Nature 2010, 468, 819–823.
- Shimizu, T.S.; Tu, Y.; Berg, H.C. A modular gradient-sensing network for chemotaxis in Escherichia coli revealed by responses to time-varying stimuli. Mol. Syst. Biol. 2010, 6, 382.
- Li, M.; Hazelbauer, G.L. Cellular Stoichiometry of the Components of the Chemotaxis Signaling Complex. J. Bacteriol. 2004, 186, 3687–3694.
- Goldbeter, A.; Koshland, D.E. An amplified sensitivity arising from covalent modification in biological systems. Proc. Natl. Acad. Sci. USA 1981, 78, 6840–6844.
- Sourjik, V.; Berg, H.C. Receptor sensitivity in bacterial chemotaxis. Proc. Natl. Acad. Sci. USA 2002, 99, 123–127.
- Colin, R.; Rosazza, C.; Vaknin, A.; Sourjik, V. Multiple sources of slow activity fluctuations in a bacterial chemosensory network. eLife 2017.
- Kamino, K.; Keegstra, J.M.; Long, J.; Emonet, T.; Shimizu, T.S. Adaptive tuning of cell sensory diversity without changes in gene expression. Sci. Adv. 2020, 6, eabc1087.
- Mattingly, H.H.; Kamino, K.; Machta, B.B.; Emonet, T. E. coli chemotaxis is information-limited. arXiv 2021, arXiv:2102.11732.
- Frank, V.; Piñas, G.E.; Cohen, H.; Parkinson, J.S.; Vaknin, A. Networked Chemoreceptors Benefit Bacterial Chemotaxis Performance. mBio 2016, 7, e1824.
- Endres, R.G.; Wingreen, N.S. Precise adaptation in bacterial chemotaxis through “assistance neighborhoods”. Proc. Natl. Acad. Sci. USA 2006, 103, 13040–13044.
- Li, M.; Hazelbauer, G.L. Adaptational assistance in clusters of bacterial chemoreceptors. Mol. Microbiol. 2005, 56, 1617–1626.
- Pontius, W.; Sneddon, M.W.; Emonet, T. Adaptation Dynamics in Densely Clustered Chemoreceptors. PLoS Comput. Biol. 2013, 9, e1003230.
- Bódi, Z.; Farkas, Z.; Nevozhay, D.; Kalapis, D.; Lázár, V.; Csörgő, B.; Nyerges, Á.; Szamecz, B.; Fekete, G.; Papp, B.; et al. Phenotypic heterogeneity promotes adaptive evolution. PLoS Biol. 2017, 15, e2000644.
- Min, T.L.; Mears, P.J.; Chubiz, L.M.; Rao, C.V.; Golding, I.; Chemla, Y.R. High-resolution, long-term characterization of bacterial motility using optical tweezers. Nat. Methods 2009, 6, 831–835.
- Min, T.L.; Mears, P.J.; Golding, I.; Chemla, Y.R. Chemotactic adaptation kinetics of individual Escherichia coli cells. Proc. Natl. Acad. Sci. USA 2012, 109, 9869–9874.
- Waite, A.J.; Frankel, N.W.; Dufour, Y.S.; Johnston, J.F.; Long, J.; Emonet, T. Non-genetic diversity modulates population performance. Mol. Syst. Biol. 2016, 12, 895.
- Dufour, Y.; Fu, X.; Hernandez-Nunez, L.; Emonet, T. Limits of Feedback Control in Bacterial Chemotaxis. PLoS Comput. Biol. 2014, 10, e1003694.
- Salek, M.M.; Carrara, F.; Fernandez, V.; Guasto, J.S.; Stocker, R. Bacterial chemotaxis in a microfluidic T-maze reveals strong phenotypic heterogeneity in chemotactic sensitivity. Nat. Commun. 2019, 10, 1–11.
- Jensen, J.L.W.V. Sur les fonctions convexes et les inégalités entre les valeurs moyennes. Acta Math. 1906, 30, 175–193.
- De Jong, I.G.; Haccou, P.; Kuipers, O.P. Bet hedging or not? A guide to proper classification of microbial survival strategies. BioEssays 2011, 33, 215–223.
- Kussell, E.; Leibler, S. Phenotypic diversity, population growth, and information in fluctuating environments. Science 2005, 309, 2075–2078.
- Shoval, O.; Sheftel, H.; Shinar, G.; Hart, Y.; Ramote, O.; Mayo, A.; Dekel, E.; Kavanagh, K.; Alon, U. Evolutionary trade-offs, Pareto optimality, and the geometry of phenotype space. Science 2012, 336, 1157–1160.
- Bhattacharjee, T.; Datta, S.S. Bacterial hopping and trapping in porous media. Nat. Commun. 2019, 10, 1–9.
- Bhattacharjee, T.; Datta, S.S. Confinement and activity regulate bacterial motion in porous media. Soft Matter 2019, 15, 9920–9930.
- Tostevin, F.; Ten Wolde, P.R. Mutual Information between Input and Output Trajectories of Biochemical Networks. Phys. Rev. Lett. 2009, 102, 218101.
- Sartori, P.; Tu, Y. Free Energy Cost of Reducing Noise while Maintaining a High Sensitivity. Phys. Rev. Lett. 2015, 115, 118102.
- Tu, Y.; Grinstein, G. How White Noise Generates Power-Law Switching in Bacterial Flagellar Motors. Phys. Rev. Lett. 2005, 94, 208101.
- Matthäus, F.; Mommer, M.S.; Curk, T.; Dobnikar, J. On the Origin and Characteristics of Noise-Induced Lévy Walks of E. Coli. PLoS ONE 2011, 6, e18623.
- Flores, M.; Shimizu, T.S.; Ten Wolde, P.R.; Tostevin, F. Signaling Noise Enhances Chemotactic Drift of E. coli. Phys. Rev. Lett. 2012, 109, 148101.
- Sneddon, M.W.; Pontius, W.; Emonet, T. Stochastic coordination of multiple actuators reduces latency and improves chemotactic response in bacteria. Proc. Natl. Acad. Sci. USA 2012, 109, 805–810.
- Huo, H.; He, R.; Zhang, R.; Yuan, J. Swimming Escherichia coli Cells Explore the Environment by Lévy Walk. Appl. Environ. Microbiol. 2021, 87, e02429-20.
- Ariel, G.; Rabani, A.; Benisty, S.; Partridge, J.D.; Harshey, R.M.; Be’er, A. Swarming bacteria migrate by Lévy Walk. Nat. Commun. 2015, 6, 8396.
- Fu, X.; Kato, S.; Long, J.; Mattingly, H.H.; He, C.; Vural, D.C.; Zucker, S.W.; Emonet, T. Spatial self-organization resolves conflicts between individuality and collective migration. Nat. Commun. 2018, 9, 1–12.
- Adler, J. Effect of Amino Acids and Oxygen on Chemotaxis in Escherichia coli. J. Bacteriol. 1966, 92, 121–129.
- Cremer, J.; Honda, T.; Tang, Y.; Ng, J.W.; Vergassola, M.; Hwa, T. Chemotaxis as a navigation strategy to boost range expansion. Nature 2019, 575, 658–663.
- Dragoš, A.; Kiesewalter, H.; Martin, M.; Hsu, C.-Y.; Hartmann, R.; Wechsler, T.; Eriksen, C.; Brix, S.; Drescher, K.; Stanley-Wall, N.; et al. Division of Labor during Biofilm Matrix Production. Curr. Biol. 2018, 28, 1903–1913.e5.
- Mattingly, H.; Emonet, T. A rule from bacteria to balance growth and expansion. Nature 2019, 575, 602–603.
- Tsoi, R.; Wu, F.; Zhang, C.; Bewick, S.; Karig, D.; You, L. Metabolic division of labor in microbial systems. Proc. Natl. Acad. Sci. USA 2018, 115, 2526–2531.
More
Information
Subjects:
Microbiology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
641
Revisions:
2 times
(View History)
Update Date:
15 Jul 2021
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No