Fibroblast growth factors (FGFs) encode a large family of growth factor proteins that activate several intracellular signaling pathways to control diverse physiological functions. The human genome encodes 22 FGFs that share a high sequence and structural homology with those of other vertebrates. FGFs orchestrate diverse biological functions by regulating cellular differentiation, proliferation, and migration. Dysregulated FGF signaling may contribute to several pathological conditions, including cancer. Notably, FGFs exhibit wide functional diversity among different vertebrates spatiotemporally. A comparative study of FGF receptor ligands and their diverse roles in vertebrates ranging from embryonic development to pathological conditions may expand our understanding of FGF.
1. Introduction
Fibroblast growth factors (FGFs) were first identified in 1973 in bovine pituitary extract [
1]. FGFs represent a family of conserved polypeptide mitogens known for their ability to promote proliferation of various cells [
2]. FGFs have pleiotropic activities that distinguish this family from other growth factors. Moreover, combined with their proliferation activity, FGFs manifest neurotrophic and angiogenic activities [
3,
4] and are involved in developmental events, including differentiation, migration, morphogenesis, and patterning [
5]. Abnormal FGF signaling causes various human diseases, such as congenital craniosynostosis, dwarfism syndrome, insulin resistance, obesity, and cancer [
6].
FGFs—present in invertebrates and vertebrates—are highly conserved in gene structure and amino acid sequence. Additionally, FGFs are the most diverse group of growth factors in vertebrates; approximately 22 members of FGFs have been identified in vertebrates, with a molecular mass of 17–34 kDa and 13–17% amino acid identity [
7]. Most FGF ligands share 28 highly conserved and six identical amino acid residues, implying similarity in the internal core [
8]. Based on sequence homology and phylogeny, vertebrate FGFs are subdivided into canonical, hormone-like, and intracellular subfamilies [
9]. Members of these subfamilies share high sequence identity and biochemical and developmental properties [
10]. Four distinct high affinity receptor tyrosine kinases mediate the effects of FGF signaling [
2].
Moreover, diversified FGF signaling requires the precise regulation of FGF activity and receptor specificity. The FGF family is extensively complex [
2], and structurally diverse FGF ligands are functionally different [
11]. However, a decade-long characterization of the structural and functional diversity within the FGF ligand family has yielded new insights on the differences in the mechanisms of action among members of the FGF family.
2. Gene Organization and Protein Length
Most
Fgf genes are dispersed throughout the vertebrate genome. The human and mouse
Fgf families comprise 22 members, and the Xenopus
Fgf family codes for approximately 19–20 FGFs [
7,
12,
13]. The location of Human
Fgf15, mouse
Fgf19, Xenopus
Fgf15,
Fgf17,
Fgf18, and
Fgf21 has not yet been identified. Moreover, evolutionarily,
Fgf15 and
Fgf19 are orthologs in vertebrates; human
Fgf19 and mouse
Fgf15 share 51% amino acid identity, and Xenopus
Fgf19 and mouse
Fgf15 share 59% identity. Additionally, a few
Fgf genes are clustered in the vertebrate genome, including
Fgf3,
Fgf4, and
Fgf19 (
Fgf15 in mice), grouped on chromosome 11 in humans and chromosome 7 in mice. However, these clustered associations of
Fgfs are common in lower vertebrates, such as Xenopus;
Fgf3, and
Fgf4, and
Fgf19 are closely linked on chromosome 4, and
Fgf1,
Fgf6,
Fgf7, and
Fgf23 are grouped on chromosome 3. Notably,
Fgf3,
Fgf4, and
Fgf19 are separated by 30 kb and 45 kb on chromosome 4 in Xenopus; however, this distance reduces to 40 kb and 10 kb in human chromosome 11. In humans and Xenopus, these gene locations indicate a conserved evolutionary pattern conferred by gene and chromosomal duplication and gene translocation.
Prototypic
Fgfs consist of three coding regions (exons), and this number is relatively conserved in humans, mice, and Xenopus. Exon 1 mainly contains the start codon (ATG); however, there are few
Fgfs (
Fgf2 and
Fgf3) where the sequence initiates from an additional 5′-transcribed sequence upstream of ATG [
14,
15]. Additionally, sub-exons are formed in some
Fgfs during the splicing process of Exon 1. The gene size of
Fgfs varies from <2 kb (in
Fgf21) to over 500 kb (in
Fgf14). Moreover, unlike other
Fgf genes, the
Fgf8 exon 1 is subdivided into four small exons in mammals [
16] followed by typical exons 2 and 3, reflecting the multifunctionality of the
Fgf8 gene. Comparing the genomic sequence of
Fgf8 genes from various species reveals that the last three exons are substantially conserved despite the upstream exons being very diverse [
16]. Based on the phylogeny chromosomal location (synteny) and homology, the
Fgf gene family in humans, mice, and Xenopus can be categorized into seven subfamilies [
17], including
Fgf1,
Fgf4,
Fgf7,
Fgf8,
Fgf9,
Fgf11, and
Fgf19/15 (
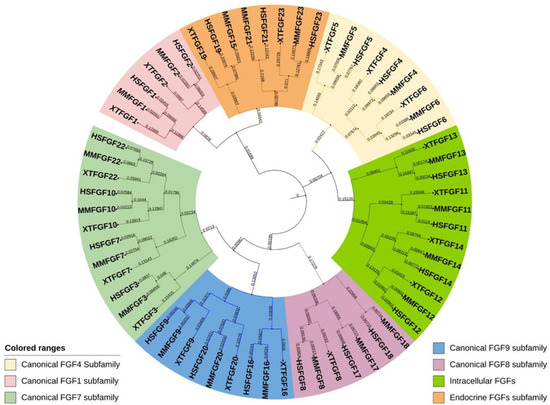
Figure 1). Phylogenetic studies suggest potential evolutionary and transformative relationships within the vertebrate gene family. Moreover, studying gene loci on chromosomes allows the evaluation of more precise evolutionary relationships within the
Fgf gene family. Lastly, the protein length of FGF is in the range of 126–268 amino acids (aa) in vertebrates, and FGFs in vertebrates are mostly of similar size; therefore, they are predicted to be similarly structured.
Figure 1. Evolutionary analysis of FGF proteins in humans, mice, and Xenopus. A circular rooted phylogenetic tree, depicting evolutionary relation between FGF proteins in Homo sapiens (HS), Mus musculus (MM), and Xenopus tropicalis (XT) was determined by using Clustal Omega and visualized by iTOL The evolutionary range among each protein is proportionate to the branch lengths and different subfamilies of FGF proteins are indicated by different color ranges.
3. Structural and Functional Diversity
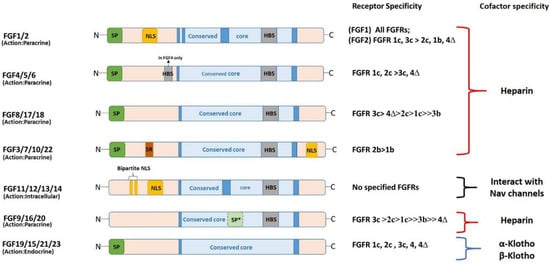
The molecular weight of FGFs in vertebrates ranges from 17–34 kDa, and the domain structure of FGF protein constitutes an internal core region of approximately 120–140 aa [
18] (
Figure 2). Within this core region, most FGFs contain a highly conserved sequence of 28 residues and 6 identical amino acids [
8]. Among the 28 highly conserved amino acid residues, 10 mediate the interaction of FGFs with their receptors (FGFR) [
19]. Moreover, the core region in most FGFs is composed of a cylindrical barrel resulting from the precise folding of 12 antiparallel β-strands. However, FGF1 and FGF2 are exceptions because their structures have a triangular array formed by the typical arrangement of four β-strands [
20]. Notably, FGF1 and FGF2—previously known as acidic and basic FGFs—were the first FGFs identified [
21]. Sharing 55% homology within their sequence, acidic and basic FGFs have acidic and basic isoelectric points of 5.6 and >9, respectively. FGF1 is a non-glycosylated polypeptide that forms a 17–18 kDa protein (155 aa in length) in most vertebrates [
22].
Figure 2. Diagrammatic representation of domain structure of FGF protein. Members of FGF family are classified based on their mode of action. N: amino terminus, SP: signal peptide, NLS: nuclear localization signal, HBS: heparin binding site, SP*: uncleaved bipartite signal sequence, C: carboxyl terminus, SR: serine rich motif.
Additionally, FGF5 (FGF3a in mice) is a precursor polypeptide containing signal and mature peptides of 17 and 251 aa, respectively, in humans. Unlike other FGFs, FGF5 is characterized by two types of glycosylation: N- and O-linked glycosylation. Lastly, FGF5 has been identified in the
Xenopus tropicalis genome [
24].
Furthermore, in the canonical subfamily, FGFs are mainly secreted ligands and are tightly bound to heparin/heparin sulfate (HS) proteoglycans (HSPGs) that regulate their receptor specificity and affinity [
29]. Members of this subfamily (FGF1, FGF4, FGF7, FGF8, and FGF9) bind to cell surface FGFRs and their cofactor protein HS to form a FGF: FGFR: HS dimer, activating in vertebrates [
30]. Notably, the FGF1 and FGF2 belong to the FGF1 subfamily.
The four major signaling pathways activated by canonical FGFs include the RAS-MAPK, phosphatidylinositol-4,5-bisphosphate 3-kinase-AKT, phospholipase Cγ/protein kinase C, and signal transducer and activator (STAT) pathways [
10]. Additionally, canonical FGFs are key regulators of mesenchymal and epithelial signaling required for organogenesis [
31].
After binding FGFR, FGF1 crosses the plasma membrane, passes through the cytosol, and reaches the nucleus [
32,
33]. Notably, FGF1 is the only FGF that can activate all splice variants of FGFR [
10], and nuclear FGF1 possibly controls the cell cycle, cell differentiation, survival, and apoptosis [
34,
35].
Phylogenically, the FGF7 subfamily includes FGF3, FGF7, FGF10, and FGF22 [
13]. However, some controversies exist regarding the inclusion of FGF3 in this subfamily, as chromosomal synteny supports its inclusion in the FGF4 subfamily [
1]. Notably, a recent study has proposed a new subfamily of FGF3 [
41]. All members of the FGF7 subfamily selectively activate splice variant IIIb of FGFR2; besides this function, FGF3 and FGF10 activate the IIIb variant of FGFR1 [
39,
40].
4. FGF Signaling in Early Development
4.1. FGF and Mesoderm Specification
Earlier investigations in the 1990s on Xenopus and other vertebrate models showed that FGF signaling is necessary for the formation of the axial (which later forms the notochord) and paraxial mesoderms (which develops into the axial skeleton, muscles, and dermis) [
60,
64]. Inhibiting FGF signaling by expressing a dominant negative form of the FGF receptor (Dn-FGFR) disrupts the notochord and somites [
60,
64,
65]. It is unclear whether FGF functions during the induction of axial and paraxial mesoderm or it is required for the maintenance of these mesodermal subtypes. Fletcher and Harland [
65] reported this dilemma in 2008, when they showed in their investigation that the induction of the paraxial mesoderm requires FGF, and axial mesoderm only requires FGF for maintenance during gastrulation. The FGF requirement for notochord development is evolutionarily conserved in vertebrates [
66].
4.2. FGF and Neural Specification
The spinal cord cells in vertebrates are derived from neuromesodermal progenitors (NMP) with neural and mesodermal features [
73,
74]. Events of spinal cord development constitute complex processes, such as neurogenesis, ventral patterning, neural crest specification, and migration, governed by the elongation of the caudal axis [
75]. Additionally, spinal cord specification involves the FGF signaling pathway as a key regulator. During chicken spinal cord specification, FGF3, FGF4, FGF8, FGF13, and FGF18 are expressed in the caudal NMP region and tissues surrounding the NMPs [
76,
77]. FGF8 and FGF4 expression in the NMP region is sustained for several days, and then declines during the last stage of somitogenesis and the cessation of axis elongation [
75].
FGF/Ras/Mapk/Ets initiate neural induction in ascidians, which are the last common ancestor of vertebrates in chordate evolution [
72,
81]. Studies in Xenopus embryos have set the foundation for the classical model (default model) of neural induction, which suggests that signals from the organizer instruct the ectoderm towards neural fate [
82]. However multiple investigations in chick embryos have established that FGF signaling is vital in early neural differentiation, challenging the default model idea [
83,
84]. FGF signaling in neuronal specification can be projected in two ways: first, as an instructive signaling where FGF activates neural genes; second, as antagonist signaling where FGF inhibits BMP signaling via smad1 phosphorylation [
12].
4.3. FGF Signaling in Metabolism and Diseases (Cancer)
FGF signaling plays a part in the development of almost every organ (including the heart, lungs, brain, urinary system, muscle, skeleton, and skin) and processes such as angiogenesis and lymphangiogenesis [
6]. Moreover, endocrine FGFs are functionally essential for metabolism and regulate the brain, kidney, liver, and adipose tissues. The dysregulation of FGF signaling leads to various genetic disorders, including cancer, chronic obstructive pulmonary disease, and chronic kidney disease.
4.3.1. FGF Signaling in Metabolism
FGF15/19, FGF21, and FGF23, which belong to the FGF19 subfamily, are endocrine hormones that regulate bile acid, fatty acid, glucose, and mineral metabolisms. Moreover, FGF19 in humans and its ortholog FGF15 are gut-derived circulating hormones that suppress hepatic bile acid via FGFR4 and the cofactor KLB complex [
6]. Additionally, FGF15/19 negatively regulates bile acid synthesis and FGF15 deletion in mice upregulates bile acid synthesis by inducing the expression of the rate-limiting and regulating enzyme cholesterol 7α-hydroxylase (CYP7A1) in the liver [
104]. However, FGF15 overexpression restricts bile acid synthesis by downregulating CYP7A1 mRNA levels [
104].
FGF15/19 suppresses liver fat storage; in one study, FGF19 transgenic mice showed low levels of lipogenic enzymes and liver triglycerides [
107]. Moreover, FGF19 blocks lipogenic enzyme expression in rat hepatocytes by inducing STAT3 signaling and suppressing peroxisome proliferator-activated receptor-γ coactivator-1β expression [
108].
FGF21 is a hormone that regulates glucose and lipid homeostasis and insulin sensitivity. FGF21 functions by binding to FGFR1c and its co-receptor protein KLB in the liver, brain, and adipose tissues [
116]. FGF21 overexpression in mice resists diet-induced obesity [
117], and FGF21 can affect weight loss, reduce plasma glucose and triglyceride levels, and boost insulin sensitivity in obese and diabetic vertebrate models without altering the calorie intake [
117,
118]. The subcutaneous administration of the FGF21 variant (LY2405319) in DIO mice decreased plasma glucose and body weight at a potency comparable to that of FGF21 [
119].
FGF23 is a regulator of phosphate metabolism and is produced mainly by the osteoblasts and osteocytes of bone tissue [
121]. Additionally, FGF23 regulates phosphate and vitamin D homeostasis in skeletal tissues [
122], and its mutations lead to low serum phosphorus levels, rickets, bone pain, osteomalacia, and short stature [
123]. Moreover, FGF23 overexpression in whole mouse, and mouse liver and osteoblasts, results in a low serum phosphate concentration and rachitic bone [
124,
125,
126].
4.3.2. FGF Signaling in Various Types of Cancer
FGFs are associated with the initiation and progression of cancers, such as multiple myeloma, urothelial carcinoma, hepatocellular carcinoma, and prostate cancer. The FGF1 expression level in several cancer types, such as breast cancer, hepatocellular carcinoma, and esophageal cancer, shows that growth factors promote tumor cell invasion and metastasis [
139,
140,
141].
FGF2 can promote the development of breast cancer cells through ligand-independent activation and the recruitment of estrogen receptor α and PRB4δ4 isoform to MYC regulatory regions [
143]. Additionally, lung cancer cells that depend on the FGF2/FGFR pathway may be prevented from proliferating using the FGF2 aptamer, which inhibits FGF2 activity [
144]. In human melanoma produced as a subcutaneous tumor model in nude mice, introducing an episomal vector encoding antisense FGF2 or FGFR1 cDNA could entirely prevent the formation of tumors by blocking angiogenesis [
145]. Targeting FGF2 to limit melanoma angiogenesis results in decisive anti-melanoma effects, which could lead to novel therapeutic approaches for patients with advanced stages of the disease.
FGF4 is expressed more frequently in germ cell cancers, particularly non-seminomas, and may target all-trans-retinoic acid to encourage the growth of malignant-cultured embryonal carcinomas [
146]. Moreover, increased FGF4 expression is linked to ovarian cancer stem-like cells’ or cancer-initiating cells’ increased capacity to initiate tumors [
147]. Furthermore, FGF5 is highly expressed in patients with breast cancer [
148], and FGF6 expression is significantly induced in metastatic liver carcinoma tissues and reduced in non-metastatic liver cancer lesion tissues [
149].
This entry is adapted from the peer-reviewed paper 10.3390/ijms24087556