1. ncRNAs and Bone Diseases
1.1. lncRNAs and SNPs in Bone Disease
Association of lncRNAs with single nucleotide polymorphism (SNP) in the coding and non-coding sequences of DNA is identified as a risk factor for the development of BMD and OP. SNP in the genomic region (1p36) has been reported as diversely linked with hip and spine BMD, and positively correlated with osteoporotic fractures [
130,
131]. In another study, Chen et al. reported that genomic variant rs6426749 (C/G) SNP at the 1p36.12 region was associated with lower BMD, and induced risk of OP. Genomic region 1p36.12 acts as an enhancer that regulates the expression of LINC00339, a lncRNA that plays a role in bone metabolism [
132,
133]. A study identified 26 specific loci in the genome that correspond to lncRNAs, efficiently associated with poor BMD and OP. In one investigation, SNP rs6894139 (T/G) in the lncRNA (MEF2C-AS1) was associated with femoral neck BMD, while SNP rs6465531 (G/A) in the lncRNA (LOC100506136) was linked with total hip BMD. Additionally, SNP rs1808124 (T/C) in BDNF-AS was associated with lower lumbar spine BMD [
130].
1.2. circRNAs and Bone Diseases
Unlike linear RNAs, circRNAs do not possess 3′ and 5′ ends and are naturally expressed as closed-loop structures. CircRNAs are endogenous RNA transcripts having limited protein-coding efficiency. Over a period, circRNAs were irrelevant byproducts without any significant biological functions. Later, thousands of circRNAs and their biogenesis were discovered [
134,
135]. circRNAs regulate proliferation, differentiation, and apoptosis in several bone pathologies, including OP, osteoarthritis (OA), osteosarcoma, and lumbar intervertebral disc degeneration [
136,
137,
138,
139]. Many circRNAs are differentially expressed; they may accelerate or repress OP. In a recent investigation, circDNAH14 (circBase ID hsa_circ_0016624) prevented OP by the regulation of BMP-2 and miR-98 sponging [
140]. In a clinical study, Liu et al. selected five samples for RNA sequencing among 40 postmenopausal osteoporosis patients (PMOP). A total of 250 differentially expressed circRNAs were estimated (64 circRNAs expressions were decreased, and 186 circRNAs were found to increase). circRNAs_0043813, 0001649, and 0005654 were upregulated, while circ_0007059, 0001204, and 0001795 were downregulated in the top six differentially expressed circRNAs. Further, Liu et al. examined circ_0007059 expression in osteoporotic samples, which were found to be reduced [
141]. About 3938 upregulated and 1505 downregulated circRNAs were shown in osteoblast differentiation [
142]. In OP, to prove the function of circRNA_0048211, Qiao et al. collected 60 samples (bone marrow) from PMOP; samples were cultured in an osteogenic induction medium. In the results, circRNA_0048211 protected OP by sponging miRNA-93-5p to regulate BMP-2 [
143]. In osteogenesis regulation, circRNAs could promote osteogenesis through upregulating FOXO1 in OP [
137]. Further, circRNAs including CDR1, CDK8, and SIPA1L1 are extensively implicated in osteogenesis differentiation [
144].
In osteoclastogenesis, circRNAs are differentially expressed. In a study conducted on mature osteoclasts, 78 miRNAs and 38 circRNAs were found to be upregulated, while 24 miRNAs and 135 circRNAs were found to be downregulated [
93]. Tumor necrosis factor-alpha (TNF-α) promotes bone resorption by osteoclast differentiation and inhibiting osteoblasts. Liu et al. examined the reduced level of circHmbox1 in TNF-α-induced osteoclast differentiation. However, circHmbox1 may inhibit RANKL-induced osteoclast differentiation by binding to miRNA-1247-5p. In OP, circRNAs have many miRNA-binding sites, function as miRNA sponges, and activate autophagy (osteoblast and osteoclast differentiation and proliferation) [
145,
146]. CircRNAs regulate several pathways, including the Wnt/β-catenin signaling pathway, BMP signaling pathway, and MAPK signaling pathway, which play a significant role in osteoporosis management. BMP2-induced osteogenesis was proved by the expression of circRNA_33287. This was upregulated in maxillary sinus membrane stem cells: circRNA_33287 downregulation inhibited several osteogenic biomarkers, such as RUNX2, ALP, and Osterix, while the upregulation exerted the opposite effect. However, circRNA_33287 is capable of inducing osteogenesis [
144,
147].
1.3. piRNAs and Bone Disease
Piwi-interacting RNAs (piRNAs) are a new subclass of ncRNAs that perform regulatory functions by explicitly interacting with Piwi proteins [
148]. piRNAs play crucial roles in the differentiation, proliferation, and maintenance of mammalian germ cells [
149,
150]. piRNAs are also expressed in somatic cells (heart, brain, bone marrow, and other tissues), some of which eliminate mRNAs (post-transcriptional level), thereby affecting disease pathogenesis [
148,
151]. Piwi proteins guide piRNA to recognize and eliminate target mRNA [
152]. In recent studies, piRNAs were expressed in the exosomes secreted by BMSCs. Exosomes are significantly expressed during the BMSCs’ osteogenic differentiation, indicating piRNAs’ contribution to osteogenesis [
153,
154]. PiR-63049 expression was shown to significantly increase in both bone tissues and plasma of PMOP and osteoporotic rats. Overexpression of piR-63049 could prevent osteoblastogenesis of BMSCs, while reduced piR-63049 expression could promote osteoblastogenesis by the Wnt2b/β-catenin signaling pathway. Additionally, in vivo knocking down of the expression of piR-63049 could attenuate bone loss by promoting bone formation. piRNA is also involved in tumor development: in patients with multiple myeloma the expression of piRNA-823 is upregulated [
148,
151]. The expression of piR-36741 is upregulated during the osteogenic differentiation of hBMSCs, while silencing of piR-36741 prominently suppresses osteogenic differentiation, resulting in reduced expression of osteogenic phenotype, osteoblast marker, and matrix mineralization. However, piR-36741 administration alleviated ovariectomy-induced osteoporosis in mice. Moreover, piR-36741 played a protective role in the osteogenic differentiation of BMSCs in mice with osteoporosis, where high expression reduced bone loss or demineralization [
154,
155].
1.4. siRNA and Bone Disease
Fire and colleagues discovered the silencing property of siRNAs in 1998 [
156]. This has become an innovative approach to downregulating the expression of the target gene, particularly knocking down the gene in vitro or in vivo. siRNA is involved in several bone-specific pathways [
157,
158]. siRNAs express enormous potential as therapeutics in managing bone disorders including OP and bone cancer. In addition, the therapeutic approach of siRNA in bone disorders can be safe and efficient. In vivo delivery of siRNA to bone-specific cells is more challenging; however, various delivery systems such as polymer-mediated delivery, peptide-based delivery, lipid-based delivery, siRNA conjugate delivery, and delivery of therapeutic siRNA in cancer cells have been developed. However, a more efficient and cell-specific delivery system is needed [
159]. Moreover, siRNA may have significant contributions to therapy. The potential ability of siRNAs is to knock down gene expression when the mRNA sequence is known. This may provide an inexpensive and efficient strategy for the management of a wide range of diseases. In bone regeneration, siRNA interferes with the expression of BMP inhibitors such as chordin and noggin, which manifests enhancing bone formation [
160,
161]. Different siRNAs target various regions of the same mRNA, with varying RNAi efficacies [
162]. Almost 58–78% of siRNAs were observed to induce silencing of genes with >50% efficiency whereas only 11–18% of siRNAs induced 90–95% silencing [
163]. To overcome siRNA delivery issues, various techniques have been developed to preserve and promote uptake by the target cells, and protect against enzymatic degradation within the cellular environment [
164].
1.5. ncRNAs and Bone Cancer
The majority of cases of osteosarcoma (OS), a high-grade primary bone tumor, are found in teens and young adults. Pathologically, this illness is marked by spindle cells and aberrant osteoid development [
165,
166]. In human malignancies, long non-coding RNAs are typically expressed abnormally and support the growth, development, and spread of tumors [
167,
168,
169,
170]. As a result, they can be used as therapeutic, diagnostic, prognostic, and predictive biomarkers [
171,
172,
173,
174]. Numerous lncRNAs with either oncogenic or tumor-suppressive functions have been reported to have differential expression in osteosarcoma. In particular, 25,733 lncRNAs were found in research by Li et al., including 403 that were constitutively elevated in 34 pathways and 798 that were downregulated in 32 pathways (by a factor of two,
p < 0.05) [
175].
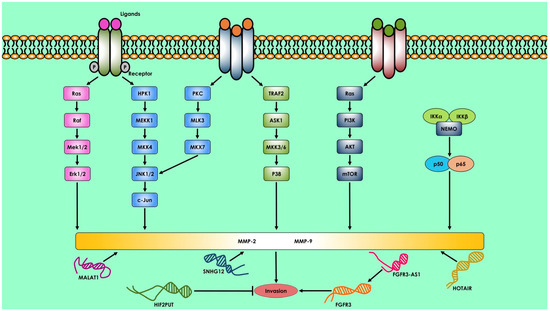
Several lncRNAs are overexpressed in osteosarcoma, and one of them is metastasis-associated lung adenocarcinoma transcript 1 (MALAT-1), a lncRNA that controls the recruitment of pre-mRNA-splicing factors to transcription sites. The level of MALAT-1 expression is closely associated with the tumor’s propensity to metastasize. Dong et al. discovered in a different investigation that MALAT-1 promotes osteosarcoma cell proliferation, migration, invasion, and lung metastasis via the PI3K/Akt pathway (
Figure 1) [
176].
Figure 1. Long non-coding RNA that regulates osteosarcoma invasion and metastasis. MALAT1, SNHG12, HOTAIR, FGFR3-AS1, and HIF2PUT are lncRNAs that govern osteosarcoma invasion and metastasis. The Erk1/2, JNK1/2, P38, PI3K/Akt, and NF-B signaling pathways control MMP-2 and MMP-9 secretion. MMP-2 and MMP-9 govern osteosarcoma cell invasion.
P50-associated COX-2 extragenic RNA (PACER), another lncRNA, is overexpressed in osteosarcoma cell lines and clinical tissues. In osteosarcoma, PACER has carcinogenic consequences by activating the COX-2 gene through the NF-B signaling cascade [
177]. LncRNA MEG3 is underexpressed in several human malignancies, including non-small-cell lung cancer, colorectal cancer, and osteosarcoma. According to gain- and loss-of-function experiments, it is controlled by the lncRNA Ewing-sarcoma-associated transcript-1 (EWSAT1). When MEG3 is downregulated in the presence of EWSAT1, osteosarcoma cells grow, invade, and migrate. As a result, the advanced clinical stage (I/II vs. III) and the existence of distant metastasis are related to lower MEG3 expression in human osteosarcoma tissue [
178,
179,
180,
181].
Another highly upregulated liver cancer lncRNA-HULC was first discovered to have an oncogenic role in human hepatocellular carcinoma. Its gene, which has a transcript length of 500 bp and connects with ribosomes, is found in the chromosomal region 6p24.3 [
182,
183]. By lowering their expression, HULC works as a sponge for many miRNAs, including miR200a-3p, miR-9, and miR107 [
184,
185]. In hepatocellular carcinoma and colorectal carcinoma cell lines, it promotes tumor cell proliferation, invasion, and angiogenesis. In osteosarcoma cell lines and tissue samples, HULC is overexpressed. This overexpression is associated with an advanced clinical stage and a low rate of overall survival in osteosarcoma patients. In osteosarcoma cell lines, HULC inhibition decreases cell growth and invasion [
186,
187,
188]. Breast, ovarian, lung, and hepatocellular carcinomas are among the malignancies in which HOTAIR is thought to be responsible for the pathogenesis [
189,
190,
191,
192]. By suppressing gene expression by histone H3K27 trimethylation and acting as a modular scaffold by binding PRC2 through the 5′ domain and LSD1/CoREST/REST complexes through the 3′ domain, it encourages the development and proliferation of tumor cells [
193,
194]. Although it is connected to other cancer forms, this molecular process in osteosarcoma is yet unclear. An intriguing case-control research in the Chinese population with 900 cases and 900 controls found that the HOTAIR gene variation rs7958904 was linked to a lower risk of osteosarcoma [
195]. The lncRNA called HOXA transcript at the distal tip (HOTTIP) is overexpressed in osteosarcoma samples and is associated with an advanced clinical stage and a significant risk of metastasis [
196]. In a number of malignant tumors, elevated expression of HOTTIP is linked to enhanced tumor cell proliferation, migration, and invasion [
196,
197,
198]. It does so through controlling RNA-binding proteins, EMT-related molecules including E-cadherin, Snail1, Slug, and others, as well as HOXA genes like HOXA13. In osteosarcoma cell lines, HOTTIP knockdown prevents cell division, migration, and invasion [
199,
200,
201].
The gene for taurine-upregulated gene 1 (TUG1), a 7.1-kb lncRNA, is found at chromosomal position 22q12.2 [
202]. To promote osteosarcoma cancer, it appears to be stimulated by p53, interacts with polycomb repressive complex 2 (PRC2), and silences specific genes implicated in the G0/G1 cell cycle arrest [
203]. In this situation, TUG1 functions as a miR-9-5p sponge and reduces POUF2F1 expression, indicating the existence of a competitive miR–lncRNA regulation network [
204]. Additionally, it stimulates the growth of osteosarcoma tumors by upregulating EZH2 through miR-144-3p. Additionally, TUG1 knockdown inhibits Wnt/-catenin pathway activation, which is overridden by EZH2 overexpression [
205]. It is interesting to note that osteosarcoma tissue clinical samples have high levels of TUG1 expression, while osteosarcoma cell line U2OS has impaired TUG1 expression, which slows cell growth and favors cell death [
202]. In osteosarcoma tissue samples, TUG1 is overexpressed, and its overexpression is linked to a poor prognosis [
206].
2. Therapeutics Approach of ncRNAs in Bone Disease
In modern medical science, ncRNAs may be the potential and efficient therapeutic option for targeting numerous bone disorders including osteoporosis, osteoarthritis, bone fractures, and bone tumors. Some important ncRNAs (lncRNA, miRNA, circRNA, piRNA, siRNA, and tRNA) and their role in managing bone disorders are discussed below.
2.1. lncRNAs in Osteoporosis and Osteoarthritis Treatment
Bone fragility and risk of fractures are common in OP patients [
207]. Huang et al. sequenced RNA (from femur subchondral tissues) and identified different gene expressions including 602 lncRNAs in patients with a femoral neck fracture and femoral head osteonecrosis. Further, data indicated that bone fractures were closely associated with specific lncRNAs. However, the differential expression of lncRNAs in fragility fractures compared with standard fractures needs to be explored [
208]. lncRNAs are crucial for proper bone healing after a bone fracture by inflammation and angiogenesis [
209]. lncRNAs also promote or suppress inflammatory pathways, implicated in bone homeostasis [
210]. The role of lncRNAs in bone fractures is still unclear. Different lncRNAs such as H19, HOTAIR, and linc-p21 are identified as altered in the bone inflammatory state (rheumatoid arthritis and osteoarthritis) (
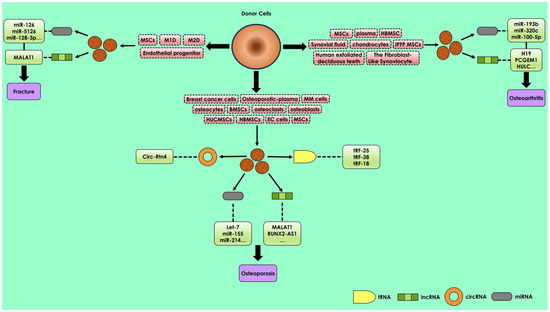
Figure 2).
Figure 2. Regulatory action of various exosomal ncRNAs including lncRNA, miRNA, circRNA, and tRNA in osteoporosis, osteoarthritis, and bone fracture repairing. The expression of exosomal ncRNA may potentially modulate bone diseases. The major source cells that contribute exosomes, which participate in osteoporosis, are BMSCs, HUCMSCs, hBMSCs, MSCs, osteocytes, osteoclasts, osteoblasts, and osteoporotic plasma; in osteoarthritis are hBMSCs, MSCs, plasma, chondrocytes, and synovial fluids; and in bone fracture repairing are MSCs, M1D, M2D, and endothelial progenitors.
Further, H19, HOTAIR, and linc-p21 express potential targets for inflammatory modulation in osteoporotic patients [
210,
211]. LncRNA HOTAIR was significantly downregulated in synoviocytes of rheumatoid arthritis patients [
212]. In rheumatoid arthritis (RA), HOTAIR overexpression reduced the secretion of IL-23 and IL-17, and decreased the number of pro-inflammatory cells (Th17), as well as diminishing levels of IL-1β, phospho-p65, and TNF-α in cartilage [
213,
214]. Moreover, lncRNA linc-p21 was found to be decreased in the blood of patients with rheumatoid arthritis, while its expression increased in human T cells [
215]. Despite this, NKILA, ANRIL, and NEAT1 also regulate NF-kB signaling, a key player in inflammatory events, contributing to proper bone healing [
216].
2.2. miRNAs in Bone Diseases and Fractures
Healing of bone fractures is the proliferative process that facilitates bone repair fracture [
217]. Initially, MSCs are recruited to the site of the fracture and differentiated as fibrocytes, osteoblasts, or chondrocytes, and these cells undergo several biomechanical stages whereby new bone is formed [
218]. The aforementioned phases are potentially regulated by miRNAs. Overexpression of miR-214-5p was reported in patients suffering from intra-articular calcaneal and/or hand fractures, while its downregulation promoted osteoblastic cell viability and resisted apoptosis [
219]. Using a mouse model with a femoral fracture, miR-186 activated the BMP signaling pathway to promote fracture healing by inhibiting
SMAD6 [
220]. Another study showed the therapeutic impact of miR-29b-3p in femoral fracture repairing: an in vivo injection of miR-29b-3p improved healing in 14 days post-fracture [
221]. The miRNAs miR-16, miR-25, miR-101, miR-19b-1, miR-92a, and miR-129-5p levels were shown to be dysregulated in bone fractures. Moreover, cells having miR-218 overexpression significantly promoted bone volume at 2 to 4 weeks post-fracture [
222,
223]. However, miRNAs regulate different cell functions but their effect in OP has not yet been well studied in well-characterized bone [
123].
2.3. Treatment of Osteoporosis by Exosomal miRNAs
Exosomal miRNAs have significant roles in the pathological process and are a clinical marker for OP diagnosis. Exosomes containing miR-21 in osteoporosis patients interfere with osteogenesis events by the potential targeting of small mothers against decapentaplegic homolog-7 (SMAD7) [
105]. In one study, MSC-derived exosomal miR-21 extracted from OP patients was expressed significantly higher than MSC-derived exosomal miR-21 extracted from healthy individuals [
224]. Song et al. showed that inhibition in the levels of exosomal miR-155 secreted by vascular endothelial cells may reverse the inhibition of osteoclast differentiation and thereby prevent bone resorption. Hence, exosomal miR-155 may have the potential to be used in treating OP [
225]. In a recent study, it was observed that exosomal miR-186 extracted from BMSCs could promote osteogenesis in OP postmenopausal women [
226]. By injecting exosomal miR-151-5p in vivo, bone reduction may be prevented [
227]. Therefore, the therapeutic impact of exosomal miRNAs in OP management has great potential. It is suggested that several miRNAs are highly expressed in exosomes, and increase or inhibit osteogenesis by regulating MSC differentiation. In OP treatments, signaling pathways, miRNAs, and related proteins have huge implications [
228].
2.4. Treatment of Osteoarthritis by Exsomal miRNAs
OA is bone degeneration in joints that causes cartilage degeneration, synovitis, chronic pain, and disability. OA is characterized by extracellular matrix (ECM) loss and cartilage destruction; treatment is focused on attenuating pain symptoms [
268]. In OP progression, OA may be treated by joint-replacement surgery. However, complete repair or regeneration of damaged articular cartilage is difficult [
269]. Exosomal miR-26a-5p derived from hBMSC are highly expressed in OA, and inhibit synovial fibroblast damage and prostaglandin-endoperoxide synthase-2, which are significant in OA treatment (
Table 1) [
248]. Existing evidence suggests that TGF-β1 regulates Sp1 through MSC-derived exosomal miR-135b, promoting chondrocyte proliferation and cartilage repair [
252]. Interestingly, using exosomal miR-92a-3p from MSCs enhances the expression of ECM genes in MSCs and promotes cartilage proliferation [
253]. The gene histone deacetylase (HDAC) is targeted by miR-193b, supporting histone (H3) acetylation and directing the metabolism of primary human chondrocytes [
261]. Interleukin-6 (IL-6)-mediated inflammation is inhibited by miR200C. In studies, exosomal miR-200C expression in synovial fluid of OA patients was found to increase by 2.5 times more than in healthy individuals [
102,
262]. Moreover, miR-4454 is associated with the inflammatory response, and miR-199b is involved in cartilage formation. Researchers have examined 50 miRNAs in the exosomes of synovial fibroblasts with differential expression [
270,
271].
Several inflammatory factors are linked to the stimulation of exosomal ncRNAs that regulate OA. A pathway, HDAC2/8, is involved in the inhibition of cartilage development through cartilage-specific genes. However, miR-95-5p in primary chondrocytes regulates cartilage development through HDAC2/8 [
254,
272]. Overexpression of miR-100-5p has been identified in stem cells that are linked with temporomandibular joint (TMJ) inflammation. Upregulation of miR expression may be associated with the occurrence and progression of OA [
102,
273].
Experimentally, a BMSC-derived exosomal miR-9-5p injection reduces inflammation and OA-like injury in the mouse. In one study, exosomal miR-9-5p targeting the syndecan-1 gene, upregulation led to an exacerbation of inflammation and OA damage [
247]. Exosome-like vesicles from chondrocytes of OA patients were shown to stimulate inflammation and increase the production of mature IL-1β by macrophages through the miR-449a-5p/ATG4B/autophagy pathway, thereby aggravating synovitis and accelerating OA progression [
274]. Chondrocytic exosomal miR-8485 stimulates cartilage differentiation in BMSCs by promoting the Wnt/β-catenin pathway [
246].
This entry is adapted from the peer-reviewed paper 10.3390/cells12081159