Halophytes and xerophytes, plants with adequate tolerance to high salinity with strong ability to survive in drought ecosystem, have been recognized for their nutritional and medicinal values owing to their comparatively higher productions of secondary metabolites, primarily the phenolics, and the flavonoids, as compared to the normal vegetation in other climatic regions. Given the consistent increases in desertification around the world, which are associated with increasing salinity, high temperature, and water scarcity, the survival of halophytes due to their secondary metabolic contents has prioritized these plant species, which have now become increasingly important for environmental protection, land reclamation, and food and animal-feed security, with their primary utility in traditional societies as sources of drugs. On the medicinal herbs front, because the fight against cancer is still ongoing, there is an urgent need for development of more efficient, safe, and novel chemotherapeutic agents, than those currently available.
- halophytes
- phenolics
- flavonoids
1. Salt-Tolerant Plants: Halophytes

2. Traditional Uses of Different Halophytes in Cancers and Cancer-Related Symptoms
3. Evaluating Anti-Cancer Activities of Halophytic Plants Extracts
| Plant | Location | Active Extract | Main Constituents | Cell Lines/In Vivo Testing, IC50 Values | Proposed Mechanism | References |
|---|---|---|---|---|---|---|
| Anabasis articulata | Saudi Arabia |
Aq. ethanolic extract |
Kaempferol 3-neohesperidosid, 6-gingerol, triterpenes, steroidal saponins, and alkaloids | Panc1 (human pancreatic cancer cell line, derived from ductal cell pancreatic carcinoma), IC50 998.5 | [70] | |
| Egypt | Methylene chloride | HePG-2 (human, hepatic carcinoma cell), IC50 6.9; HCT-116, IC50 5.5 | [162] | |||
| Arthrocnemum indicum | Tunisia | Shoot aqueous methanol extract | Gallic acid, 3-hydroxy-4′-methoxyflavone, cyanidin, chrysoeriol, quercetin, catechol, syringic acid, luteolin | Shoot extracts inhibited Caco-2 (human, colorectal adenocarcinoma cells) colon cancer cell growth in a dose-dependent manner | Cell cycle blocking at the G2/M phase | [163] |
| Arthrocnemum macrostachyum | Egypt | Methanol extract | Phenolic acids and flavonoids | In vivo anticancer effect against Ehrlich solid tumor in mice | Increased tissue necrosis and apoptosis, enhanced DNA fragmentation, upregulated cell cycle regulatory genes (Cdc2 and connexin26), and decreased TNFa levels in tumor tissues | [164] |
| Asplenium ceterach | Bulgaria | Aqueous methanol | Phenolic acids and flavonoids | A549 (human, adenocarcinoma, hypotriploid alveolar basal epithelial cells), FL, HeLa (IC50 40.48) | Strong proapoptotic potential against HeLa (human, cervical cancer cell line) |
[165] |
| Avicennia marina | Saudi Arabia | Hexane fraction | Betaine and hymecromone | HCT-116, IC50 23.7; HepG2, IC50 44.9; MCF-7, IC50 79.55 | Inhibition of cell cycle in G0/G1 and S phases in HepG2 and MCF-7 | [151] |
| Carpobrotus edulis | Portugal | Leaf methanol extract and different fractions. | β-amyrin, uvaol, oleanolic acid, monogalactosyl diacylglycerol, catechin, epicatechin, and procyanidin B5 | L5178 (mouse, T-cell lymphoma cells), and L5178 (mouse, T-cell lymphoma cells) transfected with pHa MDR1/A retrovirus | Inhibition of P-glycoprotein in MDR1-transfected mouse lymphoma cells | [166] |
| Chenopodium formosanum | Taiwan | Grain extract | Polyphenols and prebiotic dietary fiber | In vivo colon carcinogenesis induced by 1,2-dimethylhydrazine and dextran sulfate sodium in rats | Increase Bax and caspase-9 expressions; reduced TP53 and Bcl-2 expression; decreased expressions of proliferating cell nuclear antigen and cyclooxygenase-2; regulation of apoptosis-related proteins | [167,168] |
| Mesembryanthemum crystallinum | Korea | Ethanol extracts and its fractions | Phenolics and flavonoids | Inhibition of HCT116 cell growth in dose-dependent manner | Increased G2/M cell population and increased ROS levels in cells | [169] |
| Echinophora spinosa | Italy | Essential oils | p-Cymene, β-Phellandrene, β-Phellandrene, myristicin | U937, IC50 14.5–43.4 | Induced apoptosis in U937 cell line (human monocytic cell based) | [170] |
| Glaucium flavum | Tunis | Ethyl acetate extract | Isoquinoline alkaloids, kaempferol, caffeic acid, catechin hydrate, syringic acid, chlorogenic acid, isoquercitrin, and trans-hydroxycinnamic acid | MCF-7, IC50 135 | [171] | |
| Algeria | CH2Cl2 extract | MDA-MB-435, MDA-MB-231, and Hs578T (IC50 7.9–13.6) as well as in vivo tumor chorioallantoic membrane (CAM) model | Hinders angiogenesis, induction of apoptotic processes, and/or limited neovessel formation inside the tumor | [172] | ||
| Iran | Methanol extract and rich alkaloid fraction | HT-29, IC50 22.32 L; Caco-2, IC50 52.38 | [173] | |||
| Glehnia littoralis | Korea | Hexane fraction Aqueous methanol fraction |
Furanocoumarin bitter principle and polyacetylene alcohols | HT-29 (77% inhibition at 50 mg/mL extract) | Induced chromatin condensation and nuclear fragmentation, suggesting the presence of apoptotic cells; reduced mRNA expression of Bcl-2, cyclooxygenase (COX-2), and inducible nitric oxide synthase (iNOS) |
[174] |
| Limonium densiflorum | Tunisia | CHCl3 extract | Gallic acid, epigallocatechin, quercitrin, myricetin, dihydrokaempferol, isorhamnetin | A-549, IC50 29 µg/mL; DLD-1, IC50 85) | Isorhamnetin induced apoptosis through activation of peroxisome proliferator-activated receptor γ pathway in gastric cancer | [74,175] |
| Limonium bonduelli | Algeria | n-Butanol extract | Flavonoids (eriodictyol, luteolin, apigenin) and 4-hydroxy-3-methoxy benzoic acid; ethyl acetate extract of L. bonduelli and pure flavonoids, eriodictyol and luteolin | Dose-dependent growth inhibition of HT-29 and HeLa cell-lines | [176] | |
| Lotus creticus L | Portugal | Acetone extract (aerial part) Ethanol extract (fruits) |
Steroids, coumarins, tannins, and flavonoids, e.g., catechin, epicatechin, isorhamnetin, quercetin, isorhamnetin-O-hexoside, quercetin-O-hexoside, myricetin-O-hexoside | Extracts had low toxicity RAW 264.7 | [177] | |
| Lycium shawii | Saudi Arabia | Aqueous ethanol extract |
Flavonoids, 3-gluco-7-rhamnosyl quercetin, luteolin 7-O-glucoside, kaempferol-3-O-glucoside | MCF7, 194.5 µg/mL; K562, 464.9 µg/mL | Induced apoptosis and cell membrane damage due to necrosis and late apoptosis | [70] |
| Malcolmia littorea | Portugal | Polar extracts of flower and roots | Phenolic acids and flavonoids including salicylic acid and luteolin-7-O-glucoside. | HepG2 (viability 38.3%) HEK 293 cells (viability 93.1%) |
[178] | |
| Mentha piperita | China | Essential oils | Menthyl acetate, cineol, menthol, pulegone, and caryophyllene oxide | SPC-A1, IC50 10.89; K562, IC50 16.16; and SGC-7901, IC50 38.76. | [159] | |
| Pulicaria undulata | Saudi Arabia | Aqueous ethanolic extract | Flavonoids of kaempferol-, luteolin-, and quercetin-based glycosides | MCF-7, 519.2 µg/mL; K562, 1212 µg/mL; PANC-1, 1535 µg/mL |
Cell cycle arrest at the Q1 and Q2 quadrants, and necrosis in late apoptosis | [154] |
| Pulicaria crispa | Saudi Arabia | Aqueous ethanolic extract | Sterols, triterpenoids, essential oils, phenolics, and flavonoids | MDA-MB-231, IC50 180 µg/mL | Loss of cancer cell integrity, shrinkage of cytoplasm, and cell detachment | [179] |
| Reaumuria vermiculata | Tunisia | Hexane and CH2Cl2 | Myricetin, phenolics, and flavonoids | A-549, IC50 17, (hexane extract), and 23 (dichloromethane extract) | [158] | |
| Egypt | Aqueous methanol extract | Huh-7, IC50 2.4; HCT-116, IC50 1.8; MCF-7, IC50 1.3; PC-3, IC50 1.5 | [180] | |||
| Salicornia herbacea | Korea | Crude and fine polysaccharide | Polysaccharides and phenolic compounds | HT-29 | Inhibition of cyclin B1 and Cdc2 mRNAG2/M arrest | [181] |
| Salvadora persica | Saudi Arabia | Ethanol extracts of fruits | Essential oils, alkaloids, steroids, cetyl dasycarpidan-1-methanol, tetracosamethyl-cyclododecasiloxane, eicosamethyl-cyclodecasiloxane, and 1-monolinoleoylglycerol | MCF7, IC50 17.50; A2780, IC50 8.35; HT29, IC50 5.12 | [182] | |
| Salvadora persica L | Egypt | Bark petroleum ether | HepG, IC50 43.6l; MCF-7, IC50 44.3; A549, IC50 19.87 L |
[183] | ||
| Suaeda fruticosa | Pakistan | Methanol and CHCl3 extracts | Phenolics, flavonoids, saponins, fatty acids | MCF-7 (63.44% and 45.01% cell viability in methanol and CH2Cl2 at 200 μg/mL), MDA-MB-231 (77.75% and 67.22% cell viability in methanol and dichloromethane at 200 μg/mL), and DU-145 (62.83% and 25.88% cells viability in methanol and dichloromethane at 200 μg/mL) | [184] | |
| Tunisia | CH2Cl2 extract | A-549, IC50 49 ± 7; DLD-1, IC50 10 ± 1; Caco-2, IC50 140 ± 13 µg/mL; HT-29, (IC50 12 ± 14 | [157] | |||
| Saudi Arabia | Hexane extract | HCT-116, IC50 17.15; MCF-7, IC50 28.1; HepG2, IC50 33.2 | Arrest the cell cycle at the G0-G1 phase | [150] | ||
| Tamarix gallica | Tunisia | Methanolic extracts | Phenolic acids and flavonoids | Caco-2, 38% inhibition in cell growth at 100 µg/mL | Decreased DNA synthesis, arrested cell mitosis at G2/M phase; changes in the cell-cycle-associated proteins (cyclin B1, p38, Erk1/2, Chk1, and Chk2) correlated with changes in the cell cycle distribution | [185] |
| India | Protects against liver carcinogenesis initiated by diethylnitrosamine and 2-acetylaminofluorene | Restoration of cellular antioxidant enzymes, detoxifying enzymes, ODC activity, and DNA synthesis. | [161] | |||
| Zygophyllum album | Tunisia | CH2Cl2 extract | Isorhamnetin-3-O-rutinoside, quinovic acid derivatives, malvidin 3-rhamnoside, quercetin 3-sulfate | A-549, IC50 37; DLD-1, IC50 48 | Downregulation of cyclin B1 and cyclin dependent kinase; upregulation of TP53 and caspase 3 | [155] |
| Egypt | HepG2 IC50 27.74 | [186] | ||||
| Zygophyllum coccineum | Saudi Arabia | Aqueous ethanolic extract | Phenolics, flavonoids, alkaloids, quinovic acid derivatives. | MCF-7, IC50 3.47; HCT-116, IC50 3.19; HepG2, IC50 2.27 | Inhibition of human topoisomerase-IIβ | [153] |
4. Isolated-Purified Anti-Cancer Agents from Different Halophytes
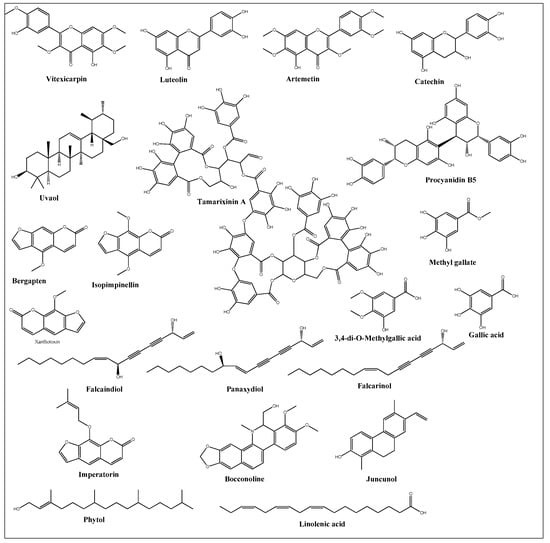
This entry is adapted from the peer-reviewed paper 10.3390/ijms24065171