The organization of eukaryotic genome in the nucleus, a double-membraned organelle separated from the cytoplasm, is highly complex and dynamic. The functional architecture of the nucleus is confined by the layers of internal and cytoplasmic elements, including chromatin organization, nuclear envelope associated proteome and transport, nuclear–cytoskeletal contacts, and the mechano-regulatory signaling cascades. The size and morphology of the nucleus could impose a significant impact on nuclear mechanics, chromatin organization, gene expression, cell functionality and disease development. The maintenance of nuclear organization during genetic or physical perturbation is crucial for the viability and lifespan of the cell. Abnormal nuclear envelope morphologies, such as invagination and blebbing, have functional implications in several human disorders, including cancer, accelerated aging, thyroid disorders, and different types of neuro-muscular diseases.
1. Introduction
The foundation of life is dependent on the functional stratification of specialized subcellular compartments. In a eukaryotic system, the nucleus forms a distinctive micro-terrain to conceal the genetic material from damaging cytoplasmic enzymes and metabolism and to provide a unique regulatory molecular framework for the genome. The spatial encapsulation of the nucleus by the lipid bilayer forms a physical and physiological intercept between cytoplasmic processes and the genome that regulates them. The construct of the nucleus is collectively furnished by a nuclear envelope along with the underlaying chromatin fiber, intermediate filaments of nucleoskeleton, nucleoplasmic subcompartments and nucleolus. These contractual components collectively impose their own effect on the rigidity, morphology and size of the nucleus [
1,
2,
3]. The nuclear shape and size are also subjected to the layers of cellular regulatory mechanisms, including C/N volume regulators, mechanobiology activated signaling cascades, macro- and micronucleophagy, etc. [
4,
5,
6,
7].
Although the nuclear size and morphology varies widely among unicellular and multicellular eukaryotes, its extent is precisely maintained in the individual cell type [
8]. However, the nucleus of same cell type may also differ among various growth phases and under different extracellular matrices. It is now understood that the nuclear, cellular or extracellular stimulants which mediate morphological alteration in the nucleus could also modulate gene expression, and therefore, the physiology of the cell [
9,
10]. The connection between nuclear structure and function has been outlined by many researchers who have categorized the nuclear pathophysiology into some broad groups, such as envelopathy, (a group of disease caused by mutation in genes encoding nuclear envelope proteins), laminopathy (diseases caused by mutations in LMNA gene) and tauopathy (a heterogeneous group of neurodegenerative diseases characterized by deposition of abnormal tau protein in the brain cells), and conferring the major responsibility for malfunctioning nuclear or cellular components to them. The structural aberrations are mostly compelled by abnormalities of nuclear envelope proteins and disorganized nucleoplasmic subcompartments, as well as hindered nucleo-cytoskeletal interactions, nuclear transport and repair mechanism. It is well-known that morphological deformations may alter cell cycle progression [
11], chromatin accessibility [
12], and the gene expression profile of a cell [
13]. Consequently, the genetic rearrangement associated with nuclear aberration could be involved with different types of malignancy, progeria syndromes, neurodegenerative diseases, neuromuscular dystrophy and many other terminal illnesses, as discussed in the following sections.
2. Contribution of Nuclear Constituents in Regulation of Nuclear Morphology
The structural components of a nucleus, such as chromosomes, nucleoplasmic compartments, nuclear envelope proteins and lipid bilayer, are the core elements involved in the regulation of nuclear morphology. Each component has a distinct functionality and approach by which they help to maintain the characteristic shape and size of the nucleus.
2.1. Nuclear Envelope Proteins
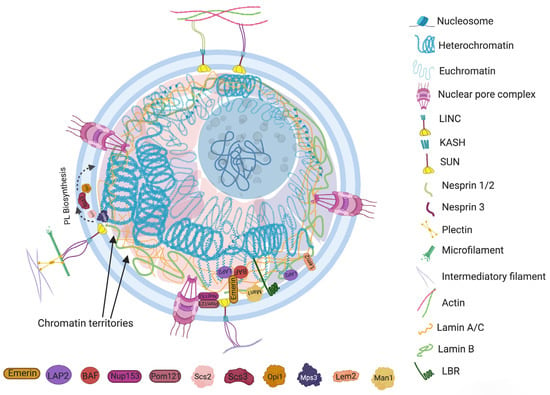
Nuclear envelopes are the structural and physiological interface between the central genomic material and cytoplasm of the cell. The double lipid membrane of the nuclear envelope originates from ER and remains in continuous contact with its network afterward. In contrast to the origin, both the outer nuclear membrane (ONM) and inner nuclear membrane (INM) of the nuclear envelope are enriched with a very distinguished set of proteomes (Figure 1) [15]. These subsets of proteins play key roles in bidirectional nucleoplasmic transportation, maintenance of nuclear architecture, cell cycle control, chromatin organization, gene regulation and DNA repair. The most complex macromolecular assemblies of the nuclear envelope are nuclear pore complexes (NPCs) [16,17,18,19]. NPCs encompass multiple subsets of more than 30 types of nuclear pore proteins called nucleoporins (Nups) [20,21]. The de novo assembly of Nups during interphase starts with the accumulation of Nups in both the outer and inner nuclear membranes, and the subsequent fusion these proteins forms the doughnut-shaped core (consisting of eight spokes arranged around a central channel) of NPC [22,23,24]. The fusion creates an energetically unfavorable and highly curved membrane that surrounds the NPC [25].
Figure 1. The constituents that contribute to regulation of morphology and characteristic organization of a common eukaryotic nucleus. The major components include the lamin network, nuclear envelope (NE), chromatin and membrane-less nuclear subcompartments. The lamin A/C and lamin B assemble around the inner nuclear membrane. Lamina and colocalized INM anchoring proteins, also known as tethering proteins (i.e., LBR, LEM, BAF, LAP, emerin, etc.), anchor at specific “lamina associated domains (LAD)” of the genome. The nuclear envelope associated components include nuclear pore complex (NPC) proteins (Nup) and the linker of nucleoskeleton and cytoskeleton (LINC) complex. Nup153 along with other membrane curvature sensing proteins (i.e., Pom) equilibrate the membrane shaping forces into the NPC assembly. Nup 153 also co-localize with Sun1 and POM121 proteins, which link the NPCs in nucleo-cytoskeletal network coupling and the mechanobiology of nuclear envelope. LINC physically connects the cytoskeletal framework to the nucleoplasmic filaments. The dynamic intermediate bridge of LINC includes INM anchored SUN domain protein and the ONM embedded KASH motif that interacts with the actin filaments, microtubule and intermediate filaments network using containing proteins, i.e., nesprin-1/2/3 and plectin, etc.; SUN domain proteins, meanwhile, bind to the NPC, lamina and chromatin using several intermediate tethering proteins. Specific INM proteins, such as Mps3, Scs2 and Opi1, contribute to lipid membrane biogenesis during morphological alteration of the nucleus. The nuclear subcompartments (Chromatin territories) are microenvironment created by the concentrate of specific proteins that contributes to the organization of different domains of chromatin fiber into the nuclear volume.
Another macromolecular assembly of NE that spans both INM and ONM is known as the linker of nucleoskeleton and cytoskeleton (LINC) complex. It physically connects the cytoskeletal framework to the nucleoplasmic filaments by forming a dynamic intermediate bridge between them. The elemental structure of LINC complex involves two transmembrane domains, ONM embedded KASH (Klarsicht, ANC-1 and SYNE homology protein) motif and INM anchored SUN (Sad1 and UNC-84 protein) domain protein. KASH motif interacts bidirectionally with SUN domain as well as with the actin filaments, microtubule and intermediate filaments network using different intermediate proteins, i.e., nesprin-1, nesprin-2, nesprin-3, dynein, kinesin and plectin, etc. In different species, at the nucleoplasmic front, various isoforms of SUN domain proteins (SUN1/2/3/4/5, Msp3, kalroid, etc.) also bind to the NPC, lamina and chromatin using several intermediate proteins [
38]. The KASH motif is a connecting link between the SUN domainand cytoskeleton. The conserved SUN domain proteins interact with lumen to carry the force aroused between cytoskeletal and nucleoskeletal network [
39]. Most importantly, the both components of LINC complex (i.e. KASH motif and SUN domain proteins) physically couple with the plasma membrane and nuclear envelope to provide a mechano-transduction signaling interface between the extracellular/cellular microenvironment and the genome [
40].
2.2. Nuclear Membrane Composition
Nuclear envelopes are one of the most functional organelles of the cell and have many simultaneous operations, including signaling, transport, genome compartmentalization, gene regulation, lipid metabolism, DNA repair and cell division, etc. These functional assortments entirely rely on the composition and physicochemical properties of the lipid membrane. The regulation of fatty acid composition of phospholipids (PL) provides specific biophysical properties, such as fluidity, rigidity or curvature to the membrane, which are required for the maintenance of the integrity and morphology of the nucleus. Interestingly, INM itself could regulate lipid composition with the help of some membrane associated proteins. It was previously noted that INM might host the lipid metabolism to expand the membrane through localized stimulation of phospholipid biosynthesis [
67]. Later, numerous proteins involved in the regulation of phospholipid biosynthesis, lipid storage and homeostasis were identified at NE [
15]. The lipid homeostasis is a complex and multifactorial mechanism that oscillates between formation of phospholipids and storage lipid using a common precursor phosphatidic acid (PA). Based on the cellular demand, PA could be converted first to diacylglycerol (DAG) and then to the storage lipid triacylglycerol (TAG); in other situations, PA could be converted into cytidine diphosphate-DAG (CDP-DAG) to form structural phospholipids.
Furthermore, in depth investigation outlines the contribution of specific INM associated proteins in lipid membrane biogenesis during morphological alteration of the nucleus. In response to the growth signals during stationary phase, a conserved PA-phosphatase Pah1 generates DAG from PA at nuclear membrane subdomain connected with storage lipid droplet. During NE growth, the activity of Pah1 is regulated by Nem1-Spo7 complex, which redirects PA towards phospholipid synthesis and membrane expansion [
68]. Many advanced studies in this line also suggest that INM localized lipid modifying proteins could also modulate nuclear morphology by transcriptional regulation of lipid synthesis genes. An interesting study by Friederichs and co-workers revealed that the nuclear morphology in budding yeast can be altered by a
mono
polar
spindle 3 (Mps3), which is lipid remodeling mechanism that uses the activity of SUN protein [
69]. The previous knowledge describes Mps3 protein as an initiator of spindle pole body (SPB) duplication and a mediator for tethering SPB to the membrane. The depletion of this protein also causes overproliferation of the inner nuclear membrane due to accumulation of abnormal amounts of polar and neutral lipids; it also inhibits the biosynthesis of sterols into the membrane [
69]. It was proposed that Mps3 promotes membrane rigidity by influencing the balance between diacyl glycerol (DAG) and phosphatidic acid (PA). Further exploration of the underlying mechanism by Ponce et al. explained that Mps3 is uniquely positioned at INM to perform along multiple pathways. Its N-terminal remains in the nucleoplasm to anchor the telomeres close to the nuclear periphery, whereas the C-terminal situated in the lumen could mediate lipid metabolism. The scholars reasoned that a link between Msp3 and Scs2 (a phospholipid biosynthesis and lipid trafficking protein) could be a possible mechanism for this behavior [
70]. Scs3 is localized at ONM and has the affinity to bind with a transcriptional corepressor of the phospholipid biosynthesis enzyme gene Opi1 [
71]. Using the connection with Scs2, Msp3 could mediate transcriptional control of lipid synthesis at the nuclear periphery (
Figure 1).
2.3. Genome Organization
The organization of the genome within the nucleus is a nonrandom process. The second level arrangement of the genome contains euchromatin and constitutive or facultative heterochromatin that gives rise to some advanced assemblies, such as chromosome loops, topological associated domains (TADs, fundamental units of three-dimensional (3D) nuclear organization), lamin associated domains (LADs, heterochromatin located adjacent to lamina), nucleolar associated domains (NADs, heterochromatin located adjacent to the nucleolus) and chromosome territories. It is also known that the nuclear arrangement of chromatin is somehow related to the morphology of the nucleus [
58,
74]. The role of chromatin in sizing and shaping the nucleus is very intricate and diverse. However, it is widely understood that chromatin contributes to nuclear morphological regulation by (i) interacting with nuclear envelope via the LAD/NAD binding domains of INM integrated proteins and (ii) altering the biophysical properties of heterochromatin.
In addition to nuclear envelope assembly, the biophysical state of constitutive and facultative heterochromatin largely influences the rigidity, shape and size of the nucleus [
2,
75]. Numerous studies have explored the role of ‘chromatin packing’ in nuclear morphology. A direct investigation was completed by Stephens et al. on chromatin decompaction of mammalian cells using histone deacetylase and histone methyltransferase inhibitors [
2]. The study showed that an increase in the ratio of euchromatin caused softer nuclei and nuclear blebbing, which was independent of the involvement of lamins. The deformation was reversed after treating the cells with histone demethylase inhibitors. It was suggested that decompacted euchromatin might be mechanically weaker than heterochromatin, or that the altered chromatin state could cause a loss of chromatin lamina connection and nuclear rigidity [
2].
2.4. Nuclear Subcompartments and Nucleolus
Nucleoplasms are among the very eventful and crowded niche of the cell that provides a common working platform to several types of heterogeneous components. It includes chromatin attached proteins and other nuclear bodies, such as nucleoli, Cajal bodies, promyelocytic leukemia (PML) bodies, speckles, paraspeckles, polycomb bodies and histone locus bodies. The consortia of nuclear bodies combine to make the nuclear matrix, which is responsible for organizing different domains of chromatin fiber into the nuclear volume [
78]. The microenvironment created by the concentration of specific proteins is referred to as membrane-less nuclear subcompartments (
Figure 1). Some components of nuclear subcompartments could also contribute to the structural organization of the nucleus. For example, Morelli and coworkers have observed that the aberrant expression of heat shock protein B2 (HSPB2), which is a nuclear subcompartment protein, in myoblast cells could cause impaired LMNA-SUN2 anchoring at the nuclear envelope, thereby disrupting NE integrity [
79]. The findings also stimulated reasonable thoughts about the impact of the nucleolus on nuclear morphology.
The nucleolus is the most prominent nuclear subcompartment and covers almost one third of the nucleoplasm’s peripheral space. Its size varies during growth, and both normal and cancer cells proliferate due to the increased demand for ribosome biogenesis [
81,
82,
83]. Almost any type of cancer exhibits abnormalities in the number and shape of nucleoli due to overactivated ribosome biosynthetic core machinery. However, it would be interesting to know whether nucleolus could have any influence on nuclear morphology in any of these conditions. The little research conducted on this topic have shown that a nucleolus has the ability to sequester the nuclear envelope to avoid nuclear morphological disruption [
84].
The direct interaction between the nuclear envelope and nucleolus was explored by some researchers. A study on breast cancer cells revealed that depletion of the nuclear envelope protein
SUN1 induced nucleolus enlargement [
85]. It is already known that INM anchored and associated proteins contribute to maintaining nuclear envelope integrity and morphology. Sharing nuclear envelope proteins to maintain nucleolar and nuclear morphology was also observed by Sen Gupta and Sengupta [
55].
2.5. Nucleus and Cytoplasmic Components
The nucleus is a largest organelle and the center of essential genetic and regulatory activities of the eukaryotic cell. Constant physiological communication among the nucleus and other cellular components, such as mitochondria, ER, vacuoles, peroxisomes, plasma membrane, lipid droplets and cytosol, maintains the cellular homeostasis [
86,
87]. Strikingly, the direct physical interconnection involving specific tethering contacts has also been recognized among the membrane-bound organelles [
88].
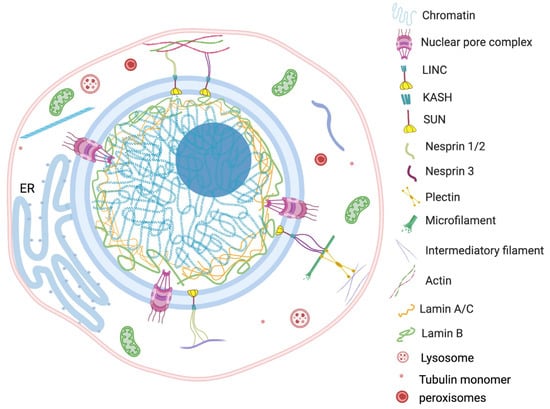
Since the nucleus is the largest and most vigorous organelle of the cell, the organization or reorganization of the cytoskeleton quickly transmits the cellular stress to the nucleus. For example, findings of Monroy-Ramírez and coworkers established that aberrant binding of tau protein and tubulin alters the radial organization of cytoskeleton to the thick ring type arrangement at peripheral and perinuclear sites [
92]. The rehabilitated nuclear–cytoskeleton assembly causes enlargement and lobulation of the nucleus followed by functional abnormalities into the cell. The externally applied tension transfers to the nucleus via the actin filament anchoring LINC complex. The direct connection between actin cytoskeleton and nuclear morphology was observed in human melanoma cells by Colón-Bolea et al. [
93]. The nuclear shape alteration in invasive melanoma cells was orchestrated by alteration in the connection between the tubulin cytoskeleton and LINC complex using a RHO GTPase (RAC1)-mediated mechanism [
93]. The concept is further corroborated by Lu et al., who demonstrated the consequence of disruption in connection between a KASH motif containing proteins and an actin network [
94]. A multivariate KASH motif containing protein, Nesprin, interacts with the actin cytoskeleton covering the outer nuclear membrane. The study revealed that Nesprin 1/ Nesprin 2 consists of a specific N-terminal actin binding domain (ABD) which is involved in actin mediated nuclear shape regulation. The overexpression of Nesprin 2 ABD leads to increase in nuclear area, but replacing it with a mini-isoform of Nesprin 2 that lacks the long rod segment produces smaller nuclei [
94]. The scholars proposed that an interchain association of Nesprin produces a basket-like protein network which has a key role in effective transduction of nuclear and cytoplasmic forces. The nuclear shape is the net outcome of external (cytoskeleton) and internal (microfilaments, lamina, genome) generated forces from opposite sides of the nuclear envelope (
Figure 2).
Figure 2. Nucleus contexture and interaction between nucleus and cytoplasmic content. The mechanical transduction of external forces affects nuclear morphology through interaction between nuclear matrix and cytoskeleton. The figure represents specific bonding between cytoplasmic macromolecules (actin, tubulin pectin, etc.) and nuclear LINC complex, lamins, SUN protein, KASH motif and nesprin protein. The physical interconnection involving the specific tethering contact of the nucleus with the membrane-bound organelles, such as ER, also plays an important role in nuclear positioning and regulation of its morphology.
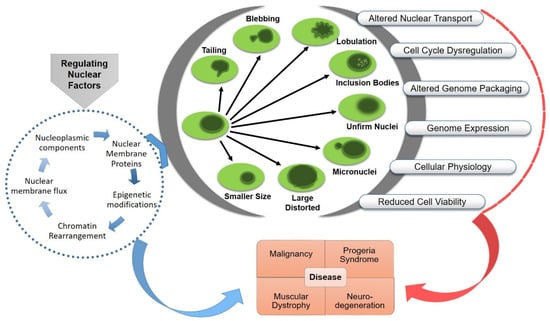
3. Functional Consequences of Nuclear Aberrations
Any morphological aberration of the nucleus could be rooted in functional abnormalities, including instability of genetic material, aneuploidy, micronuclei formation, altered gene expression and metabolic dysregulation. Nuclear pathophysiology is categorized into broad groups based on the major responsible malfunctioning component, such as envelopathy (nuclear envelope proteins that are involved in fundamental nuclear functions, such as gene transcription and DNA replication, cause human diseases through inherited or de novo mutated proteins cause human diseases, called “nuclear envelopathies”), laminopathy (diseases caused by mutations in LMNA gene, called “laminopathies”) and tauopathy (a heterogeneous group of neurodegenerative diseases characterized by abnormal metabolism of misfolded tau proteins (tau prions) which eventually results in massive loss of brain cells). Such structural aberrations affect the operational activities of the nucleus and causes devastating impact on human health, including oncogenesis, aging disorders, neuronal or muscular dystrophy or cardiomyopathy [
100,
101,
102,
103]. The pathophysiological significance of nuclear deformation has been studied exponentially in human and animal subjects, which is reflected by a tremendous number of publications in this field (
Figure 3).
Figure 3. Nuclear aberrations and functional abnormalities. Pathophysiology of related diseases and possible targets for the treatment.
The relation between nuclear deformation and progression of physiological defects are widely studied in cancer cells. In contrast to normal cells, the tumorigenic cell’s nucleus shows an unusual size and a floppy and irregular appearance due to fragmented, lobulated or deep invading outline [
104,
105]. The altered structural mechanics provide plasticity and increased invasion properties to metastatic cells; they also induce chromatin remodeling and cell cycle regulation in primary oncogenic cells [
100,
106]. Mutations in a large range of NE proteins are frequently observed in different types of cancer cells. It has been noted that the deregulation of lamin or emerin proteins could predispose mechanical distress that compromises nuclear compartmentalization and nuclear envelope integrity in cancer cells [
107,
108] and causes DNA damage in skeletal muscle cells [
109].
The rearrangement of transenvelope components, such as LINC complexes and NPCs, are required for the coordinated cell migration and attachment of invasive malignant cells [
114]. Furthermore, the altered arrangement of these nuclear envelope proteins could also modulate the genome organization that changes the nuclear mechanophysics and gene expression profile. For example, atypical Nup98 protein contributes to morphological alteration by affecting the lamina and lamina-associated polypeptides 2α (LAP2α) in leukemia cells [
115].
Lamin A farnesylation, which is key to almost all cellular defects and nuclear deformations, is also a principal prognosis component of premature aging or progeroid syndromes. Progeroid syndromes are terminal genetic disorders characterized by an accelerated aging process due to a decline in physical and physiological function at early age [
122]. Aging nucleus shows evident structural and molecular changes, including nuclear membrane lobulation and detachment, altered nuclear transport, altered genome compartmentalization and packing and an increase in transposable element transcripts and nuclear inclusions [
123,
124,
125]. The nuclear defects in progeria syndrome are caused by mutations in the LAMA gene. For example, in Hutchinson–Gilford Progeria Syndrome (HGPS), mutations in exon 11 of the
LMNA gene alters its splicing pattern and results in an in-frame deletion at C-terminus in prelamin A that produces a protein which is 50 amino acids shorter. “Progerin,” an altered prelamin A protein, interrupts the function of normal nuclear lamina at the nuclear periphery [
126]. The progerin-induced irregularities include nuclear envelope blebbing, relaxation of peripheral heterochromatin, altered epigenetic modifications and, thus, gene expression [
124,
127]. Even after correct expression of the LMNA gene, the defects in post-transcription modification of prelamin A protein may cause several premature aging diseases, including HGPS, mandibuloacral dysplasia syndrome (MAD) and restrictive dermetopathy (RD). A membrane zinc metalloprotease, ZMPSTE24, is a crucial tool for biogenesis of the lamin A scaffold protein. For the prelamin A substrate, encoded by LMNA must be farnesylated and carboxymethylated at C-terminal CAAX motif [
128]. Recessive LMNA and ZMPSTE24 mutations impede the prelamin A post-transcriptional modifications mediated by the ZMPSTE24 metalloprotease and cause cardinal nuclear morphological dysfunctions.
4. Therapeutic Approaches Targeting Nucleus in Disease and Identification of Potential Diagnostic Biomarkers
The shape of the nucleus impacts the functional status of the cell. Although the majority of cell types have either a spheroid or ovoid nucleus, different cell types can have different nuclear shapes, such as lobed, spindle shape, etc. These varied nuclear shapes have a definitive role in the transcriptional or functional activity of the cell. The human granulocytes are a good example of the need for varied nuclear shape to perform different functions. Mature neutrophils have multilobed segmented nuclei separated by thin filaments of nucleoplasm facilitating the flexibility necessary for them to pass through small gaps in the endothelium and extracellular matrix more easily. The bilobed circulating monocyte nuclei become more rounded following recruitment into tissues that further differentiate into macrophage.
The assembly of the nucleus is dynamically organized to adjust its shape and size to maintain homeostasis during different phases and needs of the cell. It is a common phenomenon of cellular functionality in which alterations in morphology happen in response to a modification in the cell’s physiological or structural environment. These morphological alterations are vital to maintain optimal functioning of the nucleus during growth and the cell’s changing needs under stress.
Nuclear morphometry plays a significant role in the histopathological and cytological diagnosis of many diseases. For instance, a 35-month follow-up study on osteosarcoma patients revealed that nuclear morphological parameters, such as area and shape, could be applied to identify which patients had a good prognosis [
161]. It was also recorded that patients with large and round tumor nuclei had better outcomes then patients with small and polymorphic nuclei. Nuclear morphological changes include alterations in size, shape, margins (grooves/molding/convolutions/thickening), shifts in chromatin pattern, enlargement of nucleoli and perinucleolar space. Morphometry and image analysis techniques are helpful to characterize the size and shape of nuclear substructures, such as nucleoli, nuclear membranes and chromatin granules. Intranuclear informatics have been developed by combined application of fluorescence microscopy, image processing and statistical analysis using specific computerized nuclear morphometric methods [
162].
Irregularities in nuclear size, shape and chromatin texture are often correlated with altered gene organization and expression in tumor cells [
11]. The remedy of such complications is completely dependent on early-stage diagnosis, when the disease is less destructive and treatment is more effective. Thereby, specific structural aberrations, including blebbing, development of nucleoplasmic reticulum, altered size and number of nucleoli and changes in nuclear rigidity have been used as important diagnostic standards to determine the type and stage of disease [
104,
163,
164].
This entry is adapted from the peer-reviewed paper 10.3390/cells12050706