Cardiovascular diseases are a major cause of death globally, with no cure to date. Many interventions have been studied and suggested, of which epigenetics and chromatin remodeling have been the most promising. Major advancements have been made in the field of chromatin remodeling, particularly for the treatment of heart failure, because of innovations in bioinformatics and gene therapy. Specifically, understanding changes to the chromatin architecture have been shown to alter cardiac disease progression via variations in genomic sequencing, targeting cardiac genes, using RNA molecules, and utilizing chromatin remodeler complexes.
- cardiovascular disease
- chromatin remodeling
- heart failure
1. Introduction
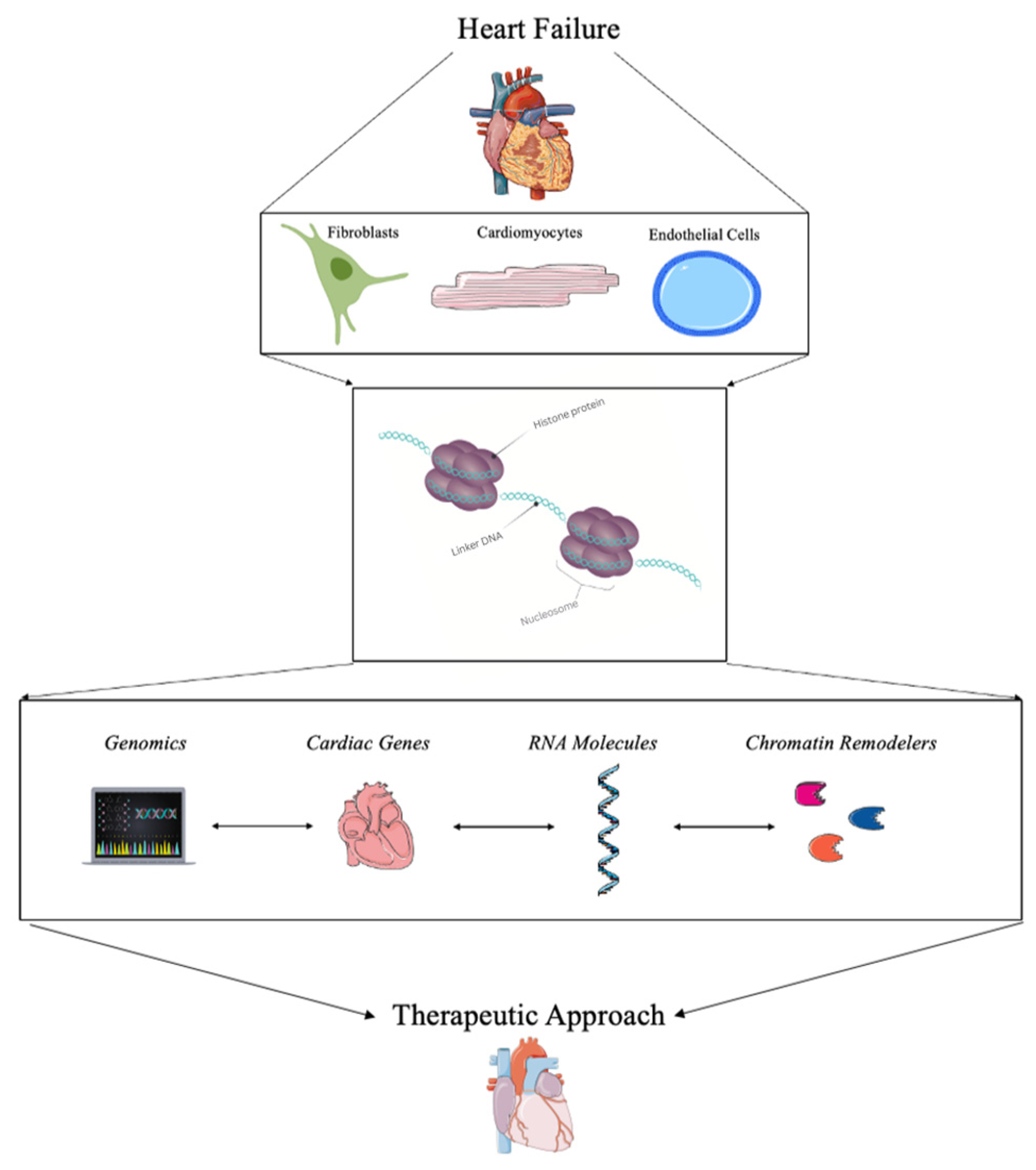
2. Current Strategies for Detecting Chromatin Remodeling in Heart Failure
3. Therapeutic Approaches Using Chromatin Remodeling in Heart Failure
3.1. Genomics and Bioinformatics
3.2. Targeting Cardiac Genes for Transcriptional Regulation
3.3. RNA Molecules
3.4. Chromatin Remodelers and Complexes
This entry is adapted from the peer-reviewed paper 10.3390/biomedicines11020579
References
- Mc Namara, K.; Alzubaidi, H.; Jackson, J.K. Cardiovascular disease as a leading cause of death: How are pharmacists getting involved? Integr. Pharm. Res. Pract. 2019, 8, 1–11.
- Cardiovascular Diseases (CVDs). Available online: https://www.who.int/news-room/fact-sheets/detail/cardiovascular-diseases-(cvds) (accessed on 17 February 2020).
- Ciumărnean, L.; Milaciu, M.V.; Negrean, V.; Orășan, O.H.; Vesa, S.C.; Sălăgean, O.; Iluţ, S.; Vlaicu, S.I. Cardiovascular Risk Factors and Physical Activity for the Prevention of Cardiovascular Diseases in the Elderly. Int. J. Environ. Res. Public Health 2022, 19, 207.
- Lifestyle Strategies for Risk Factor Reduction, Prevention and Treatme. Available online: https://www.taylorfrancis.com/chapters/edit/10.1201/9781315201108-2/lifestyle-strategies-risk-factor-reduction-prevention-treatment-cardiovascular-disease-james-rippe-theodore-angelopoulos (accessed on 6 January 2023).
- Zhao, D.; Liu, J.; Wang, M.; Zhang, X.; Zhou, M. Epidemiology of cardiovascular disease in China: Current features and implications. Nat. Rev. Cardiol. 2019, 16, 203–212.
- Savarese, G.; Lund, L.H. Global Public Health Burden of Heart Failure. Card. Fail. Rev. 2017, 3, 7–11.
- Kurmani, S.; Squire, I. Acute Heart Failure: Definition, Classification and Epidemiology. Curr. Heart Fail. Rep. 2017, 14, 385–392.
- CDC Heart Failure|cdc.gov. Centers for Disease Control and Prevention. 8 September 2020. Available online: https://www.cdc.gov/heartdisease/heart_failure.htm (accessed on 12 October 2020).
- Lippi, G.; Sanchis-Gomar, F. Global epidemiology and future trends of heart failure. AME Med. J. 2020, 5, 1–6.
- Rossignol, P.; Hernandez, A.F.; Solomon, S.D.; Zannad, F. Heart failure drug treatment. Lancet 2019, 393, 1034–1044.
- Smits, J.M.; Samuel, U.; Laufer, G. Bridging the gap in heart transplantation. Curr. Opin. Organ. Transplant. 2017, 22, 221–224.
- Rheault-Henry, M.; White, I.; Grover, D.; Atoui, R. Stem cell therapy for heart failure: Medical breakthrough, or dead end? World J. Stem Cells 2021, 13, 236–259.
- Segers, V.F.M.; Lee, R.T. Stem-cell therapy for cardiac disease. Nature 2008, 451, 937–942.
- Alexanian, M.; Padmanabhan, A.; McKinsey, T.A.; Haldar, S.M. Epigenetic therapies in heart failure. J. Mol. Cell Cardiol. 2019, 130, 197–204.
- Kim, S.Y.; Morales, C.; Gillette, T.G.; Hill, J.A. Epigenetic Regulation in Heart Failure. Curr. Opin. Cardiol. 2016, 31, 255–265.
- Chaturvedi, P.; Tyagi, S.C. Epigenetic mechanisms underlying cardiac degeneration and regeneration. Int. J. Cardiol. 2014, 173, 1–11.
- Liu, C.-F.; Tang, W.H.W. Epigenetics in Cardiac Hypertrophy and Heart Failure. J. Am. Coll. Cardiol. Basic Trans. Sci. 2019, 4, 976–993.
- McKinsey, T.A.; Vondriska, T.M.; Wang, Y. Epigenomic regulation of heart failure: Integrating histone marks, long noncoding RNAs, and chromatin architecture. F1000Res 2018, 7, F1000.
- Kimball, T.H.; Vondriska, T.M. Metabolism, Epigenetics, and Causal Inference in Heart Failure. Trends Endocrinol. Metab. 2020, 31, 181–191.
- Shi, Y.; Zhang, H.; Huang, S.; Yin, L.; Wang, F.; Luo, P.; Huang, H. Epigenetic regulation in cardiovascular disease: Mechanisms and advances in clinical trials. Sig. Transduct. Target. 2022, 7, 200.
- McGinty, R.K.; Tan, S. Histone, Nucleosome, and Chromatin Structure. In Fundamentals of Chromatin; Workman, J.L., Abmayr, S.M., Eds.; Springer: New York, NY, USA, 2014; pp. 1–28. ISBN 978-1-4614-8624-4.
- Shah, M.; He, Z.; Rauf, A.; Beikoghli Kalkhoran, S.; Heiestad, C.M.; Stensløkken, K.-O.; Parish, C.R.; Soehnlein, O.; Arjun, S.; Davidson, S.M.; et al. Extracellular histones are a target in myocardial ischaemia–reperfusion injury. Cardiovasc. Res. 2021, 118, 1115–1125.
- Gilsbach, R.; Schwaderer, M.; Preissl, S.; Grüning, B.A.; Kranzhöfer, D.; Schneider, P.; Nührenberg, T.G.; Mulero-Navarro, S.; Weichenhan, D.; Braun, C.; et al. Distinct epigenetic programs regulate cardiac myocyte development and disease in the human heart in vivo. Nat. Commun. 2018, 9, 391.
- Zhang, Q.-J.; Liu, Z.-P. Histone methylations in heart development, congenital and adult heart diseases. Epigenomics 2015, 7, 321–330.
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395.
- Lorch, Y.; Maier-Davis, B.; Kornberg, R.D. Mechanism of chromatin remodeling. Proc. Natl. Acad. Sci. USA 2010, 107, 3458–3462.
- Chromatin Remodeling in Eukaryotes|Learn Science at Scitable. Available online: http://www.nature.com/scitable/topicpage/chromatin-remodeling-in-eukaryotes-1082 (accessed on 6 January 2023).
- Pasipoularides, A. Implementing genome-driven personalized cardiology in clinical practice. J. Mol. Cell. Cardiol. 2018, 115, 142–157.
- Khomtchouk, B.B.; Tran, D.-T.; Vand, K.A.; Might, M.; Gozani, O.; Assimes, T.L. Cardioinformatics: The nexus of bioinformatics and precision cardiology. Brief. Bioinform. 2020, 21, 2031–2051.
- Hunt, C.; Montgomery, S.; Berkenpas, J.W.; Sigafoos, N.; Oakley, J.C.; Espinosa, J.; Justice, N.; Kishaba, K.; Hippe, K.; Si, D.; et al. Recent Progress of Machine Learning in Gene Therapy. Curr. Gene Ther. 2022, 22, 132–143.
- Tsompana, M.; Buck, M.J. Chromatin accessibility: A window into the genome. Epigenet. Chromatin 2014, 7, 33.
- Xu, W.; Wen, Y.; Liang, Y.; Xu, Q.; Wang, X.; Jin, W.; Chen, X. A plate-based single-cell ATAC-seq workflow for fast and robust profiling of chromatin accessibility. Nat. Protoc. 2021, 16, 4084–4107.
- Bhattacharyya, S.; Kollipara, R.K.; Orquera-Tornakian, G.; Goetsch, S.; Zhang, M.; Perry, C.; Li, B.; Shelton, J.M.; Bhakta, M.; Duan, J.; et al. Global chromatin landscapes identify candidate noncoding modifiers of cardiac rhythm. J. Clin. Investig. 2022, 133, 3.
- Jia, G.; Preussner, J.; Chen, X.; Guenther, S.; Yuan, X.; Yekelchyk, M.; Kuenne, C.; Looso, M.; Zhou, Y.; Teichmann, S.; et al. Single cell RNA-seq and ATAC-seq analysis of cardiac progenitor cell transition states and lineage settlement. Nat. Commun. 2018, 9, 4877.
- McKinsey, T.A.; Olson, E.N. Toward transcriptional therapies for the failing heart: Chemical screens to modulate genes. J. Clin. Investig. 2005, 115, 538–546.
- Centore, R.C.; Sandoval, G.J.; Soares, L.M.M.; Kadoch, C.; Chan, H.M. Mammalian SWI/SNF Chromatin Remodeling Complexes: Emerging Mechanisms and Therapeutic Strategies. Trends Genet. 2020, 36, 936–950.
- Han, P.; Chang, C.-P. Long non-coding RNA and chromatin remodeling. RNA Biol. 2015, 12, 1094–1098.
- Han, P.; Yang, J.; Shang, C.; Chang, C.-P. Chromatin Remodeling in Heart Failure. In Epigenetics in Cardiac Disease; Backs, J., McKinsey, T.A., Eds.; Cardiac and Vascullar Biology; Springer International Publishing: Cham, Switzerland, 2016; pp. 103–124. ISBN 978-3-319-41457-7.
- Han, P.; Li, W.; Lin, C.-H.; Yang, J.; Shang, C.; Nuernberg, S.T.; Jin, K.K.; Xu, W.; Lin, C.-Y.; Lin, C.-J.; et al. A long noncoding RNA protects the heart from pathological hypertrophy. Nature 2014, 514, 102–106.
- Kuppe, C.; Ramirez Flores, R.O.; Li, Z.; Hayat, S.; Levinson, R.T.; Liao, X.; Hannani, M.T.; Tanevski, J.; Wünnemann, F.; Nagai, J.S.; et al. Spatial multi-omic map of human myocardial infarction. Nature 2022, 608, 766–777.
- Chapski, D.J.; Cabaj, M.; Morselli, M.; Mason, R.J.; Soehalim, E.; Ren, S.; Pellegrini, M.; Wang, Y.; Vondriska, T.M.; Rosa-Garrido, M. Early adaptive chromatin remodeling events precede pathologic phenotypes and are reinforced in the failing heart. J. Mol. Cell. Cardiol. 2021, 160, 73–86.
- Wang, J.; Wang, Y.; Duan, Z.; Hu, W. Hypoxia-induced alterations of transcriptome and chromatin accessibility in HL-1 cells. IUBMB Life 2020, 72, 1737–1746.
- Zhou, X.; Zhang, S.; Zhao, Y.; Wang, W.; Zhang, H. A multi-omics approach to identify molecular alterations in a mouse model of heart failure. Theranostics 2022, 12, 1607–1620.
- Hall, D.D.; Spitler, K.M.; Grueter, C.E. Disruption of cardiac Med1 inhibits RNA polymerase II promoter occupancy and promotes chromatin remodeling. Am. J. Physiol.-Heart Circ. Physiol. 2019, 316, H314–H325.
- He, A.; Gu, F.; Hu, Y.; Ma, Q.; Yi Ye, L.; Akiyama, J.A.; Visel, A.; Pennacchio, L.A.; Pu, W.T. Dynamic GATA4 enhancers shape the chromatin landscape central to heart development and disease. Nat. Commun. 2014, 5, 4907.
- Jiang, W.; Xiong, Y.; Li, X.; Yang, Y. Cardiac Fibrosis: Cellular Effectors, Molecular Pathways, and Exosomal Roles. Front. Cardiovasc. Med. 2021, 8, 715258.
- Beisaw, A.; Kuenne, C.; Guenther, S.; Dallmann, J.; Wu, C.-C.; Bentsen, M.; Looso, M.; Stainier, D.Y.R. AP-1 Contributes to Chromatin Accessibility to Promote Sarcomere Disassembly and Cardiomyocyte Protrusion During Zebrafish Heart Regeneration. Circ. Res. 2020, 126, 1760–1778.
- de Pater, E.; Clijsters, L.; Marques, S.R.; Lin, Y.-F.; Garavito-Aguilar, Z.V.; Yelon, D.; Bakkers, J. Distinct phases of cardiomyocyte differentiation regulate growth of the zebrafish heart. Development 2009, 136, 1633–1641.
- Mathison, M.; Singh, V.P.; Sanagasetti, D.; Yang, L.; Pinnamaneni, J.P.; Yang, J.; Rosengart, T.K. Cardiac Reprogramming Factor Gata4 Reduces Post-Infarct Cardiac Fibrosis through Direct Repression of the Pro-Fibrotic Mediator Snail. J. Thorac. Cardiovasc. Surg. 2017, 154, 1601–1610.
- Ieda, M. Key Regulators of Cardiovascular Differentiation and Regeneration: Harnessing the Potential of Direct Reprogramming to Treat Heart Failure. J. Card. Fail. 2020, 26, 80–84.
- Yamakawa, H.; Ieda, M. Cardiac regeneration by direct reprogramming in this decade and beyond. Inflamm. Regen. 2021, 41, 20.
- McKinsey, T.A.; Foo, R.; Anene-Nzelu, C.G.; Travers, J.G.; Vagnozzi, R.J.; Weber, N.; Thum, T. Emerging epigenetic therapies of cardiac fibrosis and remodeling in heart failure: From basic mechanisms to early clinical development. Cardiovasc. Res. 2022, 118, cvac142.
- Chen, Y.; Yang, Z.; Zhao, Z.-A.; Shen, Z. Direct reprogramming of fibroblasts into cardiomyocytes. Stem Cell Res. Ther. 2017, 8, 118.
- Hu, S.; Vondriska, T.M. How chromatin stiffens fibroblasts. Curr. Opin. Physiol. 2022, 26, 100537.
- Yu, J.; Seldin, M.M.; Fu, K.; Li, S.; Lam, L.; Wang, P.; Wang, Y.; Huang, D.; Nguyen, T.L.; Wei, B.; et al. Topological Arrangement of Cardiac Fibroblasts Regulates Cellular Plasticity. Circ. Res. 2018, 123, 73–85.
- Li, Z.; Kong, X.; Zhang, Y.; Zhang, Y.; Yu, L.; Guo, J.; Xu, Y. Dual roles of chromatin remodeling protein BRG1 in angiotensin II-induced endothelial–mesenchymal transition. Cell Death Dis. 2020, 11, 549.
- Lu, P.; Ding, F.; Xiang, Y.K.; Hao, L.; Zhao, M. Noncoding RNAs in Cardiac Hypertrophy and Heart Failure. Cells 2022, 11, 777.
- Li, M.; Duan, L.; Li, Y.; Liu, B. Long noncoding RNA/circular noncoding RNA–miRNA–mRNA axes in cardiovascular diseases. Life Sci. 2019, 233, 116440.
- Gong, C.; Zhou, X.; Lai, S.; Wang, L.; Liu, J. Long Noncoding RNA/Circular RNA-miRNA-mRNA Axes in Ischemia-Reperfusion Injury. Biomed. Res. Int. 2020, 2020, 8838524.
- Chen, Y.-H.; Zhong, L.-F.; Hong, X.; Zhu, Q.-L.; Wang, S.-J.; Han, J.-B.; Huang, W.-J.; Ye, B.-Z. Integrated Analysis of circRNA-miRNA-mRNA ceRNA Network in Cardiac Hypertrophy. Front. Genet. 2022, 13, 123.
- Zhang, G.; Dou, L.; Chen, Y. Association of long-chain non-coding RNA MHRT gene single nucleotide polymorphism with risk and prognosis of chronic heart failure. Medicine 2020, 99, e19703.
- Hobuß, L.; Bär, C.; Thum, T. Long Non-coding RNAs: At the Heart of Cardiac Dysfunction? Front. Physiol. 2019, 10, 30.
- Zhang, Z.; Wan, J.; Liu, X.; Zhang, W. Strategies and technologies for exploring long noncoding RNAs in heart failure. Biomed. Pharmacother. 2020, 131, 110572.
- Hermans-Beijnsberger, S.; van Bilsen, M.; Schroen, B. Long non-coding RNAs in the failing heart and vasculature. Noncoding RNA Res. 2018, 3, 118–130.
- Tan, W.L.W.; Lim, B.T.S.; Anene-Nzelu, C.G.O.; Ackers-Johnson, M.; Dashi, A.; See, K.; Tiang, Z.; Lee, D.P.; Chua, W.W.; Luu, T.D.A.; et al. A landscape of circular RNA expression in the human heart. Cardiovasc. Res. 2017, 113, 298–309.
- Sun, C.; Ni, M.; Song, B.; Cao, L. Circulating Circular RNAs: Novel Biomarkers for Heart Failure. Front. Pharmacol. 2020, 11, 560537.
- Li, M.; Ding, W.; Tariq, M.A.; Chang, W.; Zhang, X.; Xu, W.; Hou, L.; Wang, Y.; Wang, J. A circular transcript of ncx1 gene mediates ischemic myocardial injury by targeting miR-133a-3p. Theranostics 2018, 8, 5855–5869.
- Deng, Y.; Wang, J.; Xie, G.; Zeng, X.; Li, H. Circ-HIPK3 Strengthens the Effects of Adrenaline in Heart Failure by MiR-17-3p-ADCY6 Axis. Int. J. Biol. Sci. 2019, 15, 2484–2496.
- Funamoto, M.; Sunagawa, Y.; Katanasaka, Y.; Shimizu, K.; Miyazaki, Y.; Sari, N.; Shimizu, S.; Mori, K.; Wada, H.; Hasegawa, K.; et al. Histone Acetylation Domains Are Differentially Induced during Development of Heart Failure in Dahl Salt-Sensitive Rats. Int. J. Mol. Sci. 2021, 22, 1771.
- Costantino, S.; Ambrosini, S.; Mohammed, S.A.; Gorica, E.; Akhmedov, A.; Cosentino, F.; Ruschitzka, F.; Hamdani, N.; Paneni, F. A chromatin mark by SETD7 regulates myocardial inflammation in obesity-related heart failure with preserved ejection fraction. Eur. Heart J. 2022, 43, ehac544.2883.
- Yan, F.; Chen, Z.; Cui, W. H3K9me2 regulation of BDNF expression via G9a partakes in the progression of heart failure. BMC Cardiovasc. Disord. 2022, 22, 182.
- Shi, Y.; Fan, W.; Xu, M.; Lin, X.; Zhao, W.; Yang, Z. Critical role of Znhit1 for postnatal heart function and vacuolar cardiomyopathy. JCI Insight 2022, 7, e148752.
- Sun, X.; Hota, S.K.; Zhou, Y.-Q.; Novak, S.; Miguel-Perez, D.; Christodoulou, D.; Seidman, C.E.; Seidman, J.G.; Gregorio, C.C.; Henkelman, R.M.; et al. Cardiac-enriched BAF chromatin-remodeling complex subunit Baf60c regulates gene expression programs essential for heart development and function. Biol. Open 2018, 7, bio029512.