Neurocognitive impairment (NCI) associated with HIV infection of the brain impacts a large proportion of people with HIV (PWH) regardless of antiretroviral therapy (ART). While the number of people with HIV (PWH) and severe NCI has dropped considerably with the introduction of antiretroviral therapy (ART), the sole use of ART is not sufficient to prevent or arrest NCI in many PWH. As the HIV field continues to investigate cure strategies, adjunctive therapies are greatly needed. HIV imaging, cerebrospinal fluid, and pathological studies point to the presence of continual inflammation, and the presence of HIV RNA, DNA, and proteins in the brain despite ART.
- HIV
- brain
- neuroinflammation
1. Cognitive
2. Imaging
3. Cerebrospinal Findings
4. Viral Persistence in Cerebrospinal Fluid and Viral Escape
5. Treatment
| Therapeutic | Anti-Inflammatory in CNS | AntiHIV in CNS | Other Effects | Randomized-Controlled? | Group n (Analyzed) and Summary Stats * | Outcome(s) |
|---|---|---|---|---|---|---|
| Antiretroviral | ||||||
| Maraviroc | No decrease in CSF neopterin and β2-microglobulin. Ref [92] |
NM | High EC50 in microglia in vitro. Ref [91] | ✓ | Maraviroc arm n = 9 Control arm n = 5 Time-treatment arm interaction: (p < 0.05) Effect size: (d = 0.77) and (d = 0.55) after 6 months or 12 months, respectively. |
Improved Cognition in PWH based on time-treatment arm interaction. Ref [92] |
| Cenicriviroc | NM | NM | N/A | Open label | Single arm (Cenicriviroc) n = 17 Cognitive domain of attention (p = 0.011) and working memory (p = 0.017). Decreased plasma levels of sCD163, sCD14 and neopterin (p < 0.01). |
Improved Cognition; decrease in myeloid activation markers. Ref [93] |
| Dolutegravir + Maraviroc | NM | NM | Increased CD4+ and CD8+ cell counts. | ✓ | Dual placebo n = 63 Dolutegravir and placebo n = 67 Dolutegravir and Maraviroc n = 60 cognitive testing (p > 0.10). |
No improvement in cognition in PWH + NCI. Ref [94] |
| Misc. drugs | ||||||
| Lithium | NM | NM | Lithium is well tolerated with ART. | ✓ | Placebo arm n = 31 Lithium arm n = 30 Summary Global Deficit Score-24 Weeks (p = 0.329) |
No improvement in cognition in PWH + NCI vs. placebo arm. Ref [95] |
| Selegiline transdermal system (STS) | NM | NM | Improvement in psychomotor speed in two prior pilot studies. See reference [96]. | ✓ | 3 mg/24 h STS (n = 42) 6 mg/24 h STS (n = 43) Placebo (n = 43) NPZ-8 score p = 0.35. NPZ total score p = 0.88. (Oxidative stress) CSF protein carbonyl concentration (p = 0.260) |
No improvement in cognition in STS vs placebo arm. No effect on oxidative stress. Ref [96][97] |
| Paroxetine, Fluconazole, or both | NM | NM | N/A | ✓ | Paroxetine + Fluconazole n = 11 Paroxetine + Placebo n = 11 Placebo + Fluconazole n = 9 Placebo + Placebo n = 10 Paroxetine arms improved summary NPZ-8 score (p = 0.023) |
Fluconazole did not have an observable additive effect with Paroxetine. Paroxetine in general improved cognition in PWH. Ref [98] |
| Atorvastatin | X | X | Reduction of blood lipids. | Open Label | Atorvastatin (no ART) single arm n = 7 viral load and CSF inflammatory markers (p > 0.05) Reduced cholesterol and LDL at 4–8 weeks (p < 0.01) or triglycerides at 4 weeks (p < 0.05). |
No effect on viral load or inflammatory markers in CSF. Ref [99] |
| Memantine | NM | NM | Significant increase in NAA/Cr in FWM (p= 0.040) and Parietal Cortex (p= 0.023) via MRS. | ✓ | Memantine arm n = 54 Placebo arm n = 56 NPZ-8 score p = 0.585 (week 16) |
No improvement in cognition in PWH. Ref [100] |
| Anti-inflammatory | ||||||
| Minocycline | NM | NM | Decrease in oxidative stress markers after 24 weeks. Ref [101] Decrease in various oxidative stress lipid markers (p ≤ 0.024). Ref [101] No effect on various markers of inflammation (p > 0.05) Ref [101]. |
✓ | Minocycline arm n = 26 Placebo arm n = 26 Cognitive analysis by UNP Sum criteria (p = 0.37) Ref [102]. Minocycline arm n = 8 Ref [101] Placebo arm n = 13 Ref [101] |
No improvement in cognition in PWH. Ref [102] |
6. Pathology
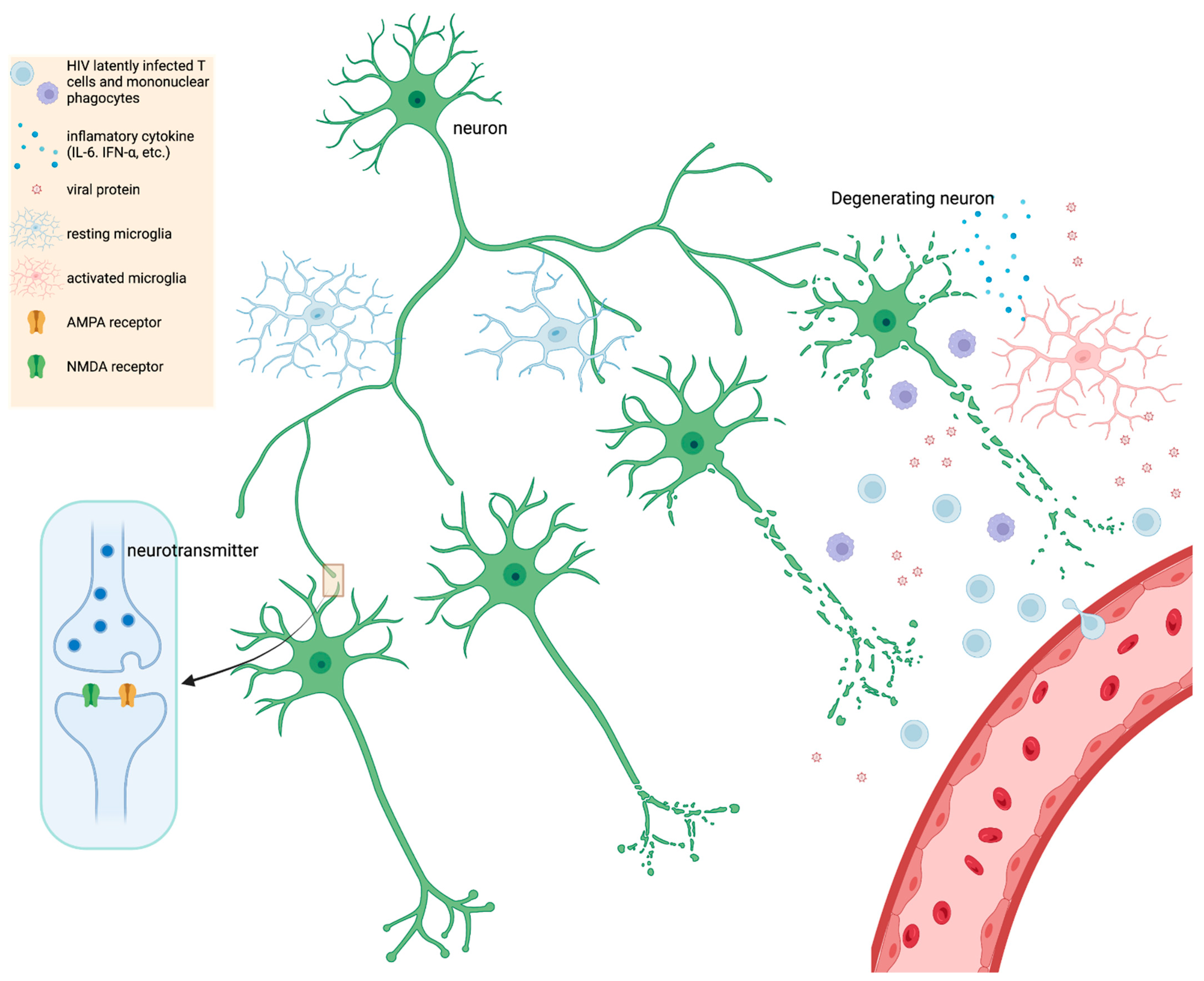
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms10112244
References
- Antinori, A.; Arendt, G.; Becker, J.T.; Brew, B.J.; Byrd, D.A.; Cherner, M.; Clifford, D.B.; Cinque, P.; Epstein, L.G.; Goodkin, K.; et al. Updated research nosology for HIV-associated neurocognitive disorders. Neurology 2007, 69, 1789–1799.
- Rumbaugh, J.A.; Tyor, W. HIV-associated neurocognitive disorders: Five new things. Neurol. Clin. Pract. 2015, 5, 224–231.
- Guaraldi, G.; Orlando, G.; Zona, S.; Menozzi, M.; Carli, F.; Garlassi, E.; Berti, A.; Rossi, E.; Roverato, A.; Palella, F. Premature age-related comorbidities among HIV-infected persons compared with the general population. Clin. Infect. Dis. 2011, 53, 1120–1126.
- Lagathu, C.; Cossarizza, A.; Béréziat, V.; Nasi, M.; Capeau, J.; Pinti, M. Basic science and pathogenesis of ageing with HIV: Potential mechanisms and biomarkers. AIDS 2017, 31 (Suppl. 2), S105–S119.
- Pathai, S.; Bajillan, H.; Landay, A.L.; High, K.P. Is HIV a model of accelerated or accentuated aging? J. Gerontol. A Biol. Sci. Med. Sci 2014, 69, 833–842.
- Lin, K.; Taylor, M.J.; Heaton, R.; Franklin, D.; Jernigan, T.; Fennema-Notestine, C.; McCutchan, A.; Atkinson, J.H.; Ellis, R.J.; McArthur, J.; et al. Effects of traumatic brain injury on cognitive functioning and cerebral metabolites in HIV-infected individuals. J. Clin. Exp. Neuropsychol. 2011, 33, 326–334.
- Heaton, R.K.; Clifford, D.B.; Franklin, D.R., Jr.; Woods, S.P.; Ake, C.; Vaida, F.; Ellis, R.J.; Letendre, S.L.; Marcotte, T.D.; Atkinson, J.H.; et al. HIV-associated neurocognitive disorders persist in the era of potent antiretroviral therapy: CHARTER Study. Neurology 2010, 75, 2087–2096.
- Bayer, C.; Tyor, W.; Tan, A.; Zola, S.; Marconi, V.; Hu, W.; Penna, S. Estimating the Prevalence and Frequency of HIV-associated Neurocognitive Disorders (HAND) in a Veteran Population. In Proceedings of the 17th Conference of the American Academy of Clinical Neuropsychology, Chicago, IL, USA, 5–8 June 2019.
- Bayer, C.; Tyor, W.; Tan, A.; Zola, S.; Hu, W.; Bott, N.; Marconi, V.; Penna, S. Utility of the Neurotrack Visual Paired Comparison (VPC) Task as a Screening Tool for HIV-associated Neurocognitive Disorders (HAND). In Proceedings of the 17th Conference of the American Academy of Clinical Neuropsychology, Chicago, IL, USA, 5–8 June 2019.
- Nightingale, S.; Winston, A. Measuring and managing cognitive impairment in HIV. AIDS 2017, 31 (Suppl. 2), S165–S172.
- Robertson, K.R.; Su, Z.; Margolis, D.M.; Krambrink, A.; Havlir, D.V.; Evans, S.; Skiest, D.J. Neurocognitive effects of treatment interruption in stable HIV-positive patients in an observational cohort. Neurology 2010, 74, 1260–1266.
- Ciavatta, V.T.; Bichler, E.K.; Speigel, I.A.; Elder, C.C.; Teng, S.L.; Tyor, W.R.; García, P.S. In vitro and Ex vivo Neurotoxic Effects of Efavirenz are Greater than Those of Other Common Antiretrovirals. Neurochem. Res. 2017, 42, 3220–3232.
- Valcour, V.; Watters, M.R.; Williams, A.E.; Sacktor, N.; McMurtray, A.; Shikuma, C. Aging exacerbates extrapyramidal motor signs in the era of highly active antiretroviral therapy. J. Neurovirol. 2008, 14, 362–367.
- Kumar, A.M.; Fernandez, J.B.; Singer, E.J.; Commins, D.; Waldrop-Valverde, D.; Ownby, R.L.; Kumar, M. Human immunodeficiency virus type 1 in the central nervous system leads to decreased dopamine in different regions of postmortem human brains. J. Neurovirol. 2009, 15, 257–274.
- Fabbiani, M.; Ciccarelli, N.; Tana, M.; Farina, S.; Baldonero, E.; Di Cristo, V.; Colafigli, M.; Tamburrini, E.; Cauda, R.; Silveri, M.C.; et al. Cardiovascular risk factors and carotid intima-media thickness are associated with lower cognitive performance in HIV-infected patients. HIV Med. 2013, 14, 136–144.
- Wendelken, L.A.; Jahanshad, N.; Rosen, H.J.; Busovaca, E.; Allen, I.; Coppola, G.; Adams, C.; Rankin, K.P.; Milanini, B.; Clifford, K.; et al. ApoE ε4 Is Associated With Cognition, Brain Integrity, and Atrophy in HIV Over Age 60. J. Acquir. Immune Defic. Syndr. 2016, 73, 426–432.
- Valcour, V.; Shikuma, C.; Shiramizu, B.; Watters, M.; Poff, P.; Selnes, O.A.; Grove, J.; Liu, Y.; Abdul-Majid, K.B.; Gartner, S.; et al. Age, apolipoprotein E4, and the risk of HIV dementia: The Hawaii Aging with HIV Cohort. J. Neuroimmunol. 2004, 157, 197–202.
- Cysique, L.A.; Soares, J.R.; Geng, G.; Scarpetta, M.; Moffat, K.; Green, M.; Brew, B.J.; Henry, R.G.; Rae, C. White matter measures are near normal in controlled HIV infection except in those with cognitive impairment and longer HIV duration. J. Neurovirol. 2017, 23, 539–547.
- Qi, Y.; Li, R.L.; Wang, Y.Y.; Wang, W.; Liu, X.Z.; Liu, J.; Li, X.; Zhang, X.D.; Yu, W.; Liu, J.J.; et al. Characteristics of Brain White Matter Microstructure in HIV Male Patients With Primary Syphilis Co-Infection. Front. Neurol. 2021, 12, 776818.
- Marra, C.M.; Deutsch, R.; Collier, A.C.; Morgello, S.; Letendre, S.; Clifford, D.; Gelman, B.; McArthur, J.; McCutchan, J.A.; Simpson, D.M.; et al. Neurocognitive impairment in HIV-infected individuals with previous syphilis. Int. J. STD AIDS 2013, 24, 351–355.
- Monick, A.J.; Joyce, M.R.; Chugh, N.; Creighton, J.A.; Morgan, O.P.; Strain, E.C.; Marvel, C.L. Characterization of basal ganglia volume changes in the context of HIV and polysubstance use. Sci. Rep. 2022, 12, 4357.
- Smail, R.C.; Brew, B.J. HIV-associated neurocognitive disorder. Handb. Clin. Neurol. 2018, 152, 75–97.
- Chang, L.; Shukla, D.K. Imaging studies of the HIV-infected brain. Handb. Clin. Neurol. 2018, 152, 229–264.
- Irollo, E.; Luchetta, J.; Ho, C.; Nash, B.; Meucci, O. Mechanisms of neuronal dysfunction in HIV-associated neurocognitive disorders. Cell Mol. Life Sci. 2021, 78, 4283–4303.
- Hassanzadeh-Behbahani, S.; Shattuck, K.F.; Bronshteyn, M.; Dawson, M.; Diaz, M.; Kumar, P.; Moore, D.J.; Ellis, R.J.; Jiang, X. Low CD4 nadir linked to widespread cortical thinning in adults living with HIV. Neuroimage Clin. 2020, 25, 102155.
- van Zoest, R.A.; Underwood, J.; De Francesco, D.; Sabin, C.A.; Cole, J.H.; Wit, F.W.; Caan, M.W.A.; Kootstra, N.A.; Fuchs, D.; Zetterberg, H.; et al. Structural Brain Abnormalities in Successfully Treated HIV Infection: Associations With Disease and Cerebrospinal Fluid Biomarkers. J. Infect. Dis. 2017, 217, 69–81.
- Chaganti, J.; Brew, B.J. MR spectroscopy in HIV associated neurocognitive disorder in the era of cART: A review. AIDS Res. Ther. 2021, 18, 65.
- Fisher, S.K.; Novak, J.E.; Agranoff, B.W. Inositol and higher inositol phosphates in neural tissues: Homeostasis, metabolism and functional significance. J. Neurochem. 2002, 82, 736–754.
- Chang, L.; Ernst, T.; Poland, R.E.; Jenden, D.J. In vivo proton magnetic resonance spectroscopy of the normal aging human brain. Life Sci. 1996, 58, 2049–2056.
- Salvan, A.M.; Vion-Dury, J.; Confort-Gouny, S.; Nicoli, F.; Lamoureux, S.; Cozzone, P.J. Brain proton magnetic resonance spectroscopy in HIV-related encephalopathy: Identification of evolving metabolic patterns in relation to dementia and therapy. AIDS Res. Hum. Retrovir. 1997, 13, 1055–1066.
- Harezlak, J.; Buchthal, S.; Taylor, M.; Schifitto, G.; Zhong, J.; Daar, E.; Alger, J.; Singer, E.; Campbell, T.; Yiannoutsos, C.; et al. Persistence of HIV-associated cognitive impairment, inflammation, and neuronal injury in era of highly active antiretroviral treatment. AIDS 2011, 25, 625–633.
- Gongvatana, A.; Harezlak, J.; Buchthal, S.; Daar, E.; Schifitto, G.; Campbell, T.; Taylor, M.; Singer, E.; Algers, J.; Zhong, J.; et al. Progressive cerebral injury in the setting of chronic HIV infection and antiretroviral therapy. J. Neurovirol. 2013, 19, 209–218.
- Young, A.C.; Yiannoutsos, C.T.; Hegde, M.; Lee, E.; Peterson, J.; Walter, R.; Price, R.W.; Meyerhoff, D.J.; Spudich, S. Cerebral metabolite changes prior to and after antiretroviral therapy in primary HIV infection. Neurology 2014, 83, 1592–1600.
- Chang, L.; Ernst, T.; Witt, M.D.; Ames, N.; Gaiefsky, M.; Miller, E. Relationships among brain metabolites, cognitive function, and viral loads in antiretroviral-naïve HIV patients. Neuroimage 2002, 17, 1638–1648.
- Meyerhoff, D.J.; Bloomer, C.; Cardenas, V.; Norman, D.; Weiner, M.W.; Fein, G. Elevated subcortical choline metabolites in cognitively and clinically asymptomatic HIV+ patients. Neurology 1999, 52, 995–1003.
- Chang, L.; Ernst, T.; St Hillaire, C.; Conant, K. Antiretroviral treatment alters relationship between MCP-1 and neurometabolites in HIV patients. Antivir. Ther. 2004, 9, 431–440.
- López-Villegas, D.; Lenkinski, R.E.; Frank, I. Biochemical changes in the frontal lobe of HIV-infected individuals detected by magnetic resonance spectroscopy. Proc. Natl. Acad. Sci. USA 1997, 94, 9854–9859.
- Coughlin, J.M.; Wang, Y.; Ma, S.; Yue, C.; Kim, P.K.; Adams, A.V.; Roosa, H.V.; Gage, K.L.; Stathis, M.; Rais, R.; et al. Regional brain distribution of translocator protein using DPA-713 PET in individuals infected with HIV. J. Neurovirol. 2014, 20, 219–232.
- Vera, J.H.; Guo, Q.; Cole, J.H.; Boasso, A.; Greathead, L.; Kelleher, P.; Rabiner, E.A.; Kalk, N.; Bishop, C.; Gunn, R.N.; et al. Neuroinflammation in treated HIV-positive individuals: A TSPO PET study. Neurology 2016, 86, 1425–1432.
- Rubin, L.H.; Sacktor, N.; Creighton, J.; Du, Y.; Endres, C.J.; Pomper, M.G.; Coughlin, J.M. Microglial activation is inversely associated with cognition in individuals living with HIV on effective antiretroviral therapy. AIDS 2018, 32, 1661–1667.
- Kim, E.J.; Yu, S.W. Translocator protein 18 kDa (TSPO): Old dogma, new mice, new structure, and new questions for neuroprotection. Neural. Regen. Res. 2015, 10, 878–880.
- Lee, Y.; Park, Y.; Nam, H.; Lee, J.W.; Yu, S.W. Translocator protein (TSPO): The new story of the old protein in neuroinflammation. BMB Rep. 2020, 53, 20–27.
- Garvey, L.J.; Pavese, N.; Politis, M.; Ramlackhansingh, A.; Brooks, D.J.; Taylor-Robinson, S.D.; Winston, A. Increased microglia activation in neurologically asymptomatic HIV-infected patients receiving effective ART. AIDS 2014, 28, 67–72.
- Boerwinkle, A.; Ances, B.M. Molecular Imaging of Neuroinflammation in HIV. J. Neuroimmune Pharmacol. 2019, 14, 9–15.
- Boerwinkle, A.H.; Strain, J.F.; Burdo, T.; Doyle, J.; Christensen, J.; Su, Y.; Wisch, J.K.; Cooley, S.A.; Vaida, F.; Smith, M.D.; et al. Comparison of -PBR28 Binding Between Persons Living With HIV and HIV-Uninfected Individuals. J. Acquir. Immune Defic. Syndr. 2020, 85, 244–251.
- Moench, T.R.; Griffin, D.E. Immunocytochemical identification and quantitation of the mononuclear cells in the cerebrospinal fluid, meninges, and brain during acute viral meningoencephalitis. J. Exp. Med. 1984, 159, 77–88.
- Rho, M.B.; Wesselingh, S.; Glass, J.D.; McArthur, J.C.; Choi, S.; Griffin, J.; Tyor, W.R. A potential role for interferon-alpha in the pathogenesis of HIV-associated dementia. Brain Behav. Immun. 1995, 9, 366–377.
- Liu, H.; Zhou, R.H.; Liu, Y.; Guo, L.; Wang, X.; Hu, W.H.; Ho, W.Z. HIV infection suppresses TLR3 activation-mediated antiviral immunity in microglia and macrophages. Immunology 2020, 160, 269–279.
- Xu, L.; Xu, Y.; Zheng, Y.; Peng, X.; Yang, Z.; Cao, Q.; Xiang, D.; Zhao, H. Differences in cytokine and chemokine profiles in cerebrospinal fluid caused by the etiology of cryptococcal meningitis and tuberculous meningitis in HIV patients. Clin. Exp. Immunol. 2021, 206, 82–90.
- Anderson, A.M.; Lennox, J.L.; Mulligan, M.M.; Loring, D.W.; Zetterberg, H.; Blennow, K.; Kessing, C.; Koneru, R.; Easley, K.; Tyor, W.R. Cerebrospinal fluid interferon alpha levels correlate with neurocognitive impairment in ambulatory HIV-Infected individuals. J. Neurovirol. 2017, 23, 106–112.
- Williams, M.E.; Stein, D.J.; Joska, J.A.; Naudé, P.J.W. Cerebrospinal fluid immune markers and HIV-associated neurocognitive impairments: A systematic review. J. Neuroimmunol. 2021, 358, 577649.
- Alammar, L.; Gama, L.; Clements, J.E. Simian immunodeficiency virus infection in the brain and lung leads to differential type I IFN signaling during acute infection. J. Immunol. 2011, 186, 4008–4018.
- Hellmuth, J.; Valcour, V.; Spudich, S. CNS reservoirs for HIV: Implications for eradication. J. Virus Erad. 2015, 1, 67–71.
- Mallard, J.; Williams, K. Correction to: An SIV macaque model of SIV and HAND: The need for adjunctive therapies in HIV that target activated monocytes and macrophages. J. Neurovirol. 2018, 24, 664.
- Eisele, E.; Siliciano, R.F. Redefining the viral reservoirs that prevent HIV-1 eradication. Immunity 2012, 37, 377–388.
- Balcom, E.F.; Roda, W.C.; Cohen, E.A.; Li, M.Y.; Power, C. HIV-1 persistence in the central nervous system: Viral and host determinants during antiretroviral therapy. Curr. Opin. Virol. 2019, 38, 54–62.
- Blankson, J.N.; Persaud, D.; Siliciano, R.F. The challenge of viral reservoirs in HIV-1 infection. Annu. Rev. Med. 2002, 53, 557–593.
- Marban, C.; Forouzanfar, F.; Ait-Ammar, A.; Fahmi, F.; El Mekdad, H.; Daouad, F.; Rohr, O.; Schwartz, C. Targeting the Brain Reservoirs: Toward an HIV Cure. Front. Immunol. 2016, 7, 397.
- Koppensteiner, H.; Brack-Werner, R.; Schindler, M. Macrophages and their relevance in Human Immunodeficiency Virus Type I infection. Retrovirology 2012, 9, 82.
- Soulet, D.; Rivest, S. Bone-marrow-derived microglia: Myth or reality? Curr. Opin. Pharmacol. 2008, 8, 508–518.
- Réu, P.; Khosravi, A.; Bernard, S.; Mold, J.E.; Salehpour, M.; Alkass, K.; Perl, S.; Tisdale, J.; Possnert, G.; Druid, H.; et al. The Lifespan and Turnover of Microglia in the Human Brain. Cell Rep. 2017, 20, 779–784.
- Roda, W.C.; Li, M.Y.; Akinwumi, M.S.; Asahchop, E.L.; Gelman, B.B.; Witwer, K.W.; Power, C. Modeling brain lentiviral infections during antiretroviral therapy in AIDS. J. Neurovirol. 2017, 23, 577–586.
- Gelman, B.B.; Lisinicchia, J.G.; Morgello, S.; Masliah, E.; Commins, D.; Achim, C.L.; Fox, H.S.; Kolson, D.L.; Grant, I.; Singer, E.; et al. Neurovirological correlation with HIV-associated neurocognitive disorders and encephalitis in a HAART-era cohort. J. Acquir. Immune Defic. Syndr. 2013, 62, 487–495.
- Valcour, V.; Chalermchai, T.; Sailasuta, N.; Marovich, M.; Lerdlum, S.; Suttichom, D.; Suwanwela, N.C.; Jagodzinski, L.; Michael, N.; Spudich, S.; et al. Central nervous system viral invasion and inflammation during acute HIV infection. J. Infect. Dis. 2012, 206, 275–282.
- Oliveira, M.F.; Chaillon, A.; Nakazawa, M.; Vargas, M.; Letendre, S.L.; Strain, M.C.; Ellis, R.J.; Morris, S.; Little, S.J.; Smith, D.M.; et al. Early Antiretroviral Therapy Is Associated with Lower HIV DNA Molecular Diversity and Lower Inflammation in Cerebrospinal Fluid but Does Not Prevent the Establishment of Compartmentalized HIV DNA Populations. PLoS Pathog. 2017, 13, e1006112.
- Dahl, V.; Peterson, J.; Fuchs, D.; Gisslen, M.; Palmer, S.; Price, R.W. Low levels of HIV-1 RNA detected in the cerebrospinal fluid after up to 10 years of suppressive therapy are associated with local immune activation. AIDS 2014, 28, 2251–2258.
- Ferretti, F.; Gisslen, M.; Cinque, P.; Price, R.W. Cerebrospinal Fluid HIV Escape from Antiretroviral Therapy. Curr. HIV/AIDS Rep. 2015, 12, 280–288.
- Nightingale, S.; Geretti, A.M.; Beloukas, A.; Fisher, M.; Winston, A.; Else, L.; Nelson, M.; Taylor, S.; Ustianowski, A.; Ainsworth, J.; et al. Discordant CSF/plasma HIV-1 RNA in patients with unexplained low-level viraemia. J. Neurovirol. 2016, 22, 852–860.
- Canestri, A.; Lescure, F.X.; Jaureguiberry, S.; Moulignier, A.; Amiel, C.; Marcelin, A.G.; Peytavin, G.; Tubiana, R.; Pialoux, G.; Katlama, C. Discordance between cerebral spinal fluid and plasma HIV replication in patients with neurological symptoms who are receiving suppressive antiretroviral therapy. Clin. Infect. Dis. 2010, 50, 773–778.
- Peluso, M.J.; Ferretti, F.; Peterson, J.; Lee, E.; Fuchs, D.; Boschini, A.; Gisslén, M.; Angoff, N.; Price, R.W.; Cinque, P.; et al. Cerebrospinal fluid HIV escape associated with progressive neurologic dysfunction in patients on antiretroviral therapy with well controlled plasma viral load. AIDS 2012, 26, 1765–1774.
- Kugathasan, R.; Collier, D.A.; Haddow, L.J.; El Bouzidi, K.; Edwards, S.G.; Cartledge, J.D.; Miller, R.F.; Gupta, R.K. Diffuse White Matter Signal Abnormalities on Magnetic Resonance Imaging Are Associated With Human Immunodeficiency Virus Type 1 Viral Escape in the Central Nervous System Among Patients With Neurological Symptoms. Clin. Infect. Dis. 2017, 64, 1059–1065.
- Mastrangelo, A.; Turrini, F.; de Zan, V.; Caccia, R.; Gerevini, S.; Cinque, P. Symptomatic cerebrospinal fluid escape. AIDS 2019, 33 (Suppl. 2), S159–S169.
- Levy, D.N.; Refaeli, Y.; MacGregor, R.R.; Weiner, D.B. Serum Vpr regulates productive infection and latency of human immunodeficiency virus type 1. Proc. Natl. Acad. Sci. USA 1994, 91, 10873–10877.
- Mamik, M.K.; Hui, E.; Branton, W.G.; McKenzie, B.A.; Chisholm, J.; Cohen, E.A.; Power, C. HIV-1 Viral Protein R Activates NLRP3 Inflammasome in Microglia: Implications for HIV-1 Associated Neuroinflammation. J. Neuroimmune Pharmacol. 2017, 12, 233–248.
- Evering, T.H.; Kamau, E.; St Bernard, L.; Farmer, C.B.; Kong, X.P.; Markowitz, M. Single genome analysis reveals genetic characteristics of Neuroadaptation across HIV-1 envelope. Retrovirology 2014, 11, 65.
- Henderson, L.J.; Johnson, T.P.; Smith, B.R.; Reoma, L.B.; Santamaria, U.A.; Bachani, M.; Demarino, C.; Barclay, R.A.; Snow, J.; Sacktor, N.; et al. Presence of Tat and transactivation response element in spinal fluid despite antiretroviral therapy. AIDS 2019, 33 (Suppl. 2), S145–S157.
- Jones, G.J.; Barsby, N.L.; Cohen, E.A.; Holden, J.; Harris, K.; Dickie, P.; Jhamandas, J.; Power, C. HIV-1 Vpr causes neuronal apoptosis and in vivo neurodegeneration. J. Neurosci. 2007, 27, 3703–3711.
- Kitayama, H.; Miura, Y.; Ando, Y.; Hoshino, S.; Ishizaka, Y.; Koyanagi, Y. Human immunodeficiency virus type 1 Vpr inhibits axonal outgrowth through induction of mitochondrial dysfunction. J. Virol. 2008, 82, 2528–2542.
- Zou, W.; Kim, B.O.; Zhou, B.Y.; Liu, Y.; Messing, A.; He, J.J. Protection against human immunodeficiency virus type 1 Tat neurotoxicity by Ginkgo biloba extract EGb 761 involving glial fibrillary acidic protein. Am. J. Pathol. 2007, 171, 1923–1935.
- Kim, B.O.; Liu, Y.; Ruan, Y.; Xu, Z.C.; Schantz, L.; He, J.J. Neuropathologies in transgenic mice expressing human immunodeficiency virus type 1 Tat protein under the regulation of the astrocyte-specific glial fibrillary acidic protein promoter and doxycycline. Am. J. Pathol. 2003, 162, 1693–1707.
- Bachis, A.; Wenzel, E.; Boelk, A.; Becker, J.; Mocchetti, I. The neurotrophin receptor p75 mediates gp120-induced loss of synaptic spines in aging mice. Neurobiol. Aging 2016, 46, 160–168.
- Toggas, S.M.; Masliah, E.; Rockenstein, E.M.; Rall, G.F.; Abraham, C.R.; Mucke, L. Central nervous system damage produced by expression of the HIV-1 coat protein gp120 in transgenic mice. Nature 1994, 367, 188–193.
- Thaney, V.E.; O’Neill, A.M.; Hoefer, M.M.; Maung, R.; Sanchez, A.B.; Kaul, M. IFNβ Protects Neurons from Damage in a Murine Model of HIV-1 Associated Brain Injury. Sci. Rep. 2017, 7, 46514.
- Ditiatkovski, M.; Mukhamedova, N.; Dragoljevic, D.; Hoang, A.; Low, H.; Pushkarsky, T.; Fu, Y.; Carmichael, I.; Hill, A.F.; Murphy, A.J.; et al. Modification of lipid rafts by extracellular vesicles carrying HIV-1 protein Nef induces redistribution of amyloid precursor protein and Tau, causing neuronal dysfunction. J. Biol. Chem. 2020, 295, 13377–13392.
- Rosenthal, J.; Tyor, W. Aging, comorbidities, and the importance of finding biomarkers for HIV-associated neurocognitive disorders. J. Neurovirol. 2019, 25, 673–685.
- Anderson, A.M.; Jang, J.H.; Easley, K.A.; Fuchs, D.; Gisslen, M.; Zetterberg, H.; Blennow, K.; Ellis, R.J.; Franklin, D.; Heaton, R.K.; et al. Cognitive and Neuronal Link With Inflammation: A Longitudinal Study in People With and Without HIV Infection. J. Acquir. Immune Defic. Syndr. 2020, 85, 617–625.
- Robertson, K.R.; Robertson, W.T.; Ford, S.; Watson, D.; Fiscus, S.; Harp, A.G.; Hall, C.D. Highly active antiretroviral therapy improves neurocognitive functioning. J. Acquir. Immune Defic. Syndr. 2004, 36, 562–566.
- Schmitt, F.A.; Bigley, J.W.; McKinnis, R.; Logue, P.E.; Evans, R.W.; Drucker, J.L. Neuropsychological outcome of zidovudine (AZT) treatment of patients with AIDS and AIDS-related complex. N. Engl. J. Med. 1988, 319, 1573–1578.
- Letendre, S.; Marquie-Beck, J.; Capparelli, E.; Best, B.; Clifford, D.; Collier, A.C.; Gelman, B.B.; McArthur, J.C.; McCutchan, J.A.; Morgello, S.; et al. Validation of the CNS Penetration-Effectiveness rank for quantifying antiretroviral penetration into the central nervous system. Arch. Neurol. 2008, 65, 65–70.
- Letendre, S.L.; Ellis, R.J.; Ances, B.M.; McCutchan, J.A. Neurologic complications of HIV disease and their treatment. Top HIV Med. 2010, 18, 45–55.
- Asahchop, E.L.; Meziane, O.; Mamik, M.K.; Chan, W.F.; Branton, W.G.; Resch, L.; Gill, M.J.; Haddad, E.; Guimond, J.V.; Wainberg, M.A.; et al. Reduced antiretroviral drug efficacy and concentration in HIV-infected microglia contributes to viral persistence in brain. Retrovirology 2017, 14, 47.
- Gates, T.M.; Cysique, L.A.; Siefried, K.J.; Chaganti, J.; Moffat, K.J.; Brew, B.J. Maraviroc-intensified combined antiretroviral therapy improves cognition in virally suppressed HIV-associated neurocognitive disorder. AIDS 2016, 30, 591–600.
- D’Antoni, M.L.; Paul, R.H.; Mitchell, B.I.; Kohorn, L.; Fischer, L.; Lefebvre, E.; Seyedkazemi, S.; Nakamoto, B.K.; Walker, M.; Kallianpur, K.J.; et al. Improved Cognitive Performance and Reduced Monocyte Activation in Virally Suppressed Chronic HIV After Dual CCR2 and CCR5 Antagonism. J. Acquir. Immune Defic. Syndr. 2018, 79, 108–116.
- Letendre, S.L.; Roa, J.; Chen, H.; McKhann, A.; Marra, C.M.; Daar, E.S.; Hunt, P.W.; Campbell, T.; Swaminathan, S.; Ha, B.; et al. ACTG A5324: A Randomized Trial of ART Intensification for Cognitive Impairment in PWH. In Proceedings of the Conference on Retroviruses and Opportunistic Infections, Virtual, 12–16 February 2022.
- Decloedt, E.H.; Freeman, C.; Howells, F.; Casson-Crook, M.; Lesosky, M.; Koutsilieri, E.; Lovestone, S.; Maartens, G.; Joska, J.A. Moderate to severe HIV-associated neurocognitive impairment: A randomized placebo-controlled trial of lithium. Medicine 2016, 95, e5401.
- Schifitto, G.; Yiannoutsos, C.T.; Ernst, T.; Navia, B.A.; Nath, A.; Sacktor, N.; Anderson, C.; Marra, C.M.; Clifford, D.B. Selegiline and oxidative stress in HIV-associated cognitive impairment. Neurology 2009, 73, 1975–1981.
- Schifitto, G.; Zhang, J.; Evans, S.R.; Sacktor, N.; Simpson, D.; Millar, L.L.; Hung, V.L.; Miller, E.N.; Smith, E.; Ellis, R.J.; et al. A multicenter trial of selegiline transdermal system for HIV-associated cognitive impairment. Neurology 2007, 69, 1314–1321.
- Sacktor, N.; Skolasky, R.L.; Moxley, R.; Wang, S.; Mielke, M.M.; Munro, C.; Steiner, J.; Nath, A.; Haughey, N.; McArthur, J. Paroxetine and fluconazole therapy for HIV-associated neurocognitive impairment: Results from a double-blind, placebo-controlled trial. J. Neurovirol. 2018, 24, 16–27.
- Probasco, J.C.; Spudich, S.S.; Critchfield, J.; Lee, E.; Lollo, N.; Deeks, S.G.; Price, R.W. Failure of atorvastatin to modulate CSF HIV-1 infection: Results of a pilot study. Neurology 2008, 71, 521–524.
- Schifitto, G.; Navia, B.A.; Yiannoutsos, C.T.; Marra, C.M.; Chang, L.; Ernst, T.; Jarvik, J.G.; Miller, E.N.; Singer, E.J.; Ellis, R.J.; et al. Memantine and HIV-associated cognitive impairment: A neuropsychological and proton magnetic resonance spectroscopy study. AIDS 2007, 21, 1877–1886.
- Sacktor, N.; Miyahara, S.; Evans, S.; Schifitto, G.; Cohen, B.; Haughey, N.; Drewes, J.L.; Graham, D.; Zink, M.C.; Anderson, C.; et al. Impact of minocycline on cerebrospinal fluid markers of oxidative stress, neuronal injury, and inflammation in HIV-seropositive individuals with cognitive impairment. J. Neurovirol. 2014, 20, 620–626.
- Nakasujja, N.; Miyahara, S.; Evans, S.; Lee, A.; Musisi, S.; Katabira, E.; Robertson, K.; Ronald, A.; Clifford, D.B.; Sacktor, N. Randomized trial of minocycline in the treatment of HIV-associated cognitive impairment. Neurology 2013, 80, 196–202.
- Ndhlovu, L.C.; D’Antoni, M.L.; Ananworanich, J.; Byron, M.M.; Chalermchai, T.; Sithinamsuwan, P.; Tipsuk, S.; Ho, E.; Slike, B.M.; Schuetz, A.; et al. Loss of CCR2 expressing non-classical monocytes are associated with cognitive impairment in antiretroviral therapy-naïve HIV-infected Thais. J. Neuroimmunol. 2015, 288, 25–33.
- Lin, S.P.; Calcagno, A.; Letendre, S.L.; Ma, Q. Clinical Treatment Options and Randomized Clinical Trials for Neurocognitive Complications of HIV Infection: Combination Antiretroviral Therapy, Central Nervous System Penetration Effectiveness, and Adjuvants. Curr. Top Behav. Neurosci. 2021, 50, 517–545.
- Steiner, J.P.; Bachani, M.; Wolfson-Stofko, B.; Lee, M.H.; Wang, T.; Li, G.; Li, W.; Strayer, D.; Haughey, N.J.; Nath, A. Interaction of paroxetine with mitochondrial proteins mediates neuroprotection. Neurotherapeutics 2015, 12, 200–216.
- Meulendyke, K.A.; Queen, S.E.; Engle, E.L.; Shirk, E.N.; Liu, J.; Steiner, J.P.; Nath, A.; Tarwater, P.M.; Graham, D.R.; Mankowski, J.L.; et al. Combination fluconazole/paroxetine treatment is neuroprotective despite ongoing neuroinflammation and viral replication in an SIV model of HIV neurological disease. J. Neurovirol. 2014, 20, 591–602.
- Ashraf, T.; Jiang, W.; Hoque, M.T.; Henderson, J.; Wu, C.; Bendayan, R. Role of anti-inflammatory compounds in human immunodeficiency virus-1 glycoprotein120-mediated brain inflammation. J. Neuroinflammation 2014, 11, 91.
- Yadav, A.; Betts, M.R.; Collman, R.G. Statin modulation of monocyte phenotype and function: Implications for HIV-1-associated neurocognitive disorders. J. Neurovirol. 2016, 22, 584–596.
- Bandaru, V.V.; Mielke, M.M.; Sacktor, N.; McArthur, J.C.; Grant, I.; Letendre, S.; Chang, L.; Wojna, V.; Pardo, C.; Calabresi, P.; et al. A lipid storage-like disorder contributes to cognitive decline in HIV-infected subjects. Neurology 2013, 81, 1492–1499.
- Saylor, D.; Molsberry, S.A.; Seaberg, E.C.; Cheng, Y.; Levine, A.; Martin, E.; Munro, C.; Palella, F.; Becker, J.; Sacktor, N. Statin Use and Cognitive Performance in the Multicenter Aids Cohort Study. In Proceedings of the Conference on Retroviruses and Opportunistic Infections, Virtual, 6–11 March 2021.
- Marconi, V.C.; Moser, C.; Gavegnano, C.; Deeks, S.G.; Lederman, M.M.; Overton, E.T.; Tsibris, A.; Hunt, P.W.; Kantor, A.; Sekaly, R.P.; et al. Randomized Trial of Ruxolitinib in Antiretroviral-Treated Adults With Human Immunodeficiency Virus. Clin. Infect. Dis. 2022, 74, 95–104.
- Bastard, J.P.; Soulié, C.; Fellahi, S.; Haïm-Boukobza, S.; Simon, A.; Katlama, C.; Calvez, V.; Marcelin, A.G.; Capeau, J. Circulating interleukin-6 levels correlate with residual HIV viraemia and markers of immune dysfunction in treatment-controlled HIV-infected patients. Antivir. Ther. 2012, 17, 915–919.
- Lichtfuss, G.F.; Cheng, W.J.; Farsakoglu, Y.; Paukovics, G.; Rajasuriar, R.; Velayudham, P.; Kramski, M.; Hearps, A.C.; Cameron, P.U.; Lewin, S.R.; et al. Virologically suppressed HIV patients show activation of NK cells and persistent innate immune activation. J. Immunol. 2012, 189, 1491–1499.
- Kamat, A.; Misra, V.; Cassol, E.; Ancuta, P.; Yan, Z.; Li, C.; Morgello, S.; Gabuzda, D. A plasma biomarker signature of immune activation in HIV patients on antiretroviral therapy. PLoS ONE 2012, 7, e30881.
- Lederman, M.M.; Funderburg, N.T.; Sekaly, R.P.; Klatt, N.R.; Hunt, P.W. Residual immune dysregulation syndrome in treated HIV infection. Adv. Immunol. 2013, 119, 51–83.
- Gavegnano, C.; Brehm, J.H.; Dupuy, F.P.; Talla, A.; Ribeiro, S.P.; Kulpa, D.A.; Cameron, C.; Santos, S.; Hurwitz, S.J.; Marconi, V.C.; et al. Novel mechanisms to inhibit HIV reservoir seeding using Jak inhibitors. PLoS Pathog. 2017, 13, e1006740.
- Vier, J.; Groth, M.; Sochalska, M.; Kirschnek, S. The anti-apoptotic Bcl-2 family protein A1/Bfl-1 regulates neutrophil survival and homeostasis and is controlled via PI3K and JAK/STAT signaling. Cell Death Dis. 2016, 7, e2103.
- Renault, T.T.; Chipuk, J.E. Getting away with murder: How does the BCL-2 family of proteins kill with immunity? Ann. N. Y. Acad. Sci. 2013, 1285, 59–79.
- Chetoui, N.; Boisvert, M.; Gendron, S.; Aoudjit, F. Interleukin-7 promotes the survival of human CD4+ effector/memory T cells by up-regulating Bcl-2 proteins and activating the JAK/STAT signalling pathway. Immunology 2010, 130, 418–426.
- Gavegnano, C.; Haile, W.B.; Hurwitz, S.; Tao, S.; Jiang, Y.; Schinazi, R.F.; Tyor, W.R. Baricitinib reverses HIV-associated neurocognitive disorders in a SCID mouse model and reservoir seeding in vitro. J. Neuroinflammation 2019, 16, 182.
- Marconi, V.C.; Ramanan, A.V.; de Bono, S.; Kartman, C.E.; Krishnan, V.; Liao, R.; Piruzeli, M.L.B.; Goldman, J.D.; Alatorre-Alexander, J.; de Cassia Pellegrini, R.; et al. Efficacy and safety of baricitinib for the treatment of hospitalised adults with COVID-19 (COV-BARRIER): A randomised, double-blind, parallel-group, placebo-controlled phase 3 trial. Lancet Respir. Med. 2021, 9, 1407–1418.
- Titanji, B.K.; Farley, M.M.; Mehta, A.; Connor-Schuler, R.; Moanna, A.; Cribbs, S.K.; O’Shea, J.; DeSilva, K.; Chan, B.; Edwards, A.; et al. Use of Baricitinib in Patients With Moderate to Severe Coronavirus Disease 2019. Clin. Infect. Dis. 2021, 72, 1247–1250.
- Bronte, V.; Ugel, S.; Tinazzi, E.; Vella, A.; De Sanctis, F.; Canè, S.; Batani, V.; Trovato, R.; Fiore, A.; Petrova, V.; et al. Baricitinib restrains the immune dysregulation in patients with severe COVID-19. J. Clin. Investig. 2020, 130, 6409–6416.
- Kubo, S.; Nakayamada, S.; Sakata, K.; Kitanaga, Y.; Ma, X.; Lee, S.; Ishii, A.; Yamagata, K.; Nakano, K.; Tanaka, Y. Janus Kinase Inhibitor Baricitinib Modulates Human Innate and Adaptive Immune System. Front. Immunol. 2018, 9, 1510.
- Gelman, B.B. Neuropathology of HAND With Suppressive Antiretroviral Therapy: Encephalitis and Neurodegeneration Reconsidered. Curr. HIV/AIDS Rep. 2015, 12, 272–279.
- Kolson, D.L.; Sabnekar, P.; Baybis, M.; Crino, P.B. Gene expression in TUNEL-positive neurons in human immunodeficiency virus-infected brain. J. Neurovirol. 2004, 10 (Suppl. 1), 102–107.
- Masliah, E.; Ge, N.; Morey, M.; DeTeresa, R.; Terry, R.D.; Wiley, C.A. Cortical dendritic pathology in human immunodeficiency virus encephalitis. Lab Investig. 1992, 66, 285–291.
- Masliah, E.; Heaton, R.K.; Marcotte, T.D.; Ellis, R.J.; Wiley, C.A.; Mallory, M.; Achim, C.L.; McCutchan, J.A.; Nelson, J.A.; Atkinson, J.H.; et al. Dendritic injury is a pathological substrate for human immunodeficiency virus-related cognitive disorders. HNRC Group. The HIV Neurobehavioral Research Center. Ann. Neurol. 1997, 42, 963–972.
- Adle-Biassette, H.; Chretien, F.; Wingertsmann, L.; Hery, C.; Ereau, T.; Scaravilli, F.; Tardieu, M.; Gray, F. Neuronal apoptosis does not correlate with dementia in HIV infection but is related to microglial activation and axonal damage. Neuropathol. Appl. Neurobiol. 1999, 25, 123–133.
- Ellis, R.; Langford, D.; Masliah, E. HIV and antiretroviral therapy in the brain: Neuronal injury and repair. Nat. Rev. Neurosci. 2007, 8, 33–44.
- Weis, S.; Haug, H.; Budka, H. Neuronal damage in the cerebral cortex of AIDS brains: A morphometric study. Acta Neuropathol. 1993, 85, 185–189.
- Carroll, A.; Brew, B. HIV-associated neurocognitive disorders: Recent advances in pathogenesis, biomarkers, and treatment. F1000Res 2017, 6, 312.
- Manji, H.; Jäger, H.R.; Winston, A. HIV, dementia and antiretroviral drugs: 30 years of an epidemic. J. Neurol. Neurosurg. Psychiatry 2013, 84, 1126–1137.
- Spudich, S.; Gisslen, M.; Hagberg, L.; Lee, E.; Liegler, T.; Brew, B.; Fuchs, D.; Tambussi, G.; Cinque, P.; Hecht, F.M.; et al. Central nervous system immune activation characterizes primary human immunodeficiency virus 1 infection even in participants with minimal cerebrospinal fluid viral burden. J. Infect. Dis. 2011, 204, 753–760.
- Green, M.V.; Raybuck, J.D.; Zhang, X.; Wu, M.M.; Thayer, S.A. Scaling Synapses in the Presence of HIV. Neurochem. Res. 2019, 44, 234–246.
- Brown, J.C.; Higgins, E.S.; George, M.S. Synaptic Plasticity 101: The Story of the AMPA Receptor for the Brain Stimulation Practitioner. Neuromodulation 2021.
- Koneru, R.; Bimonte-Nelson, H.; Ciavatta, V.; Haile, W.; Elmore, K.; Ward, J.; Maroun, L.; Tyor, W.R. Reversing interferon-alpha neurotoxicity in a HIV-associated neurocognitive disorder mouse model. AIDS 2018, 32, 1403–1411.
- Cornell, J.; Salinas, S.; Huang, H.Y.; Zhou, M. Microglia regulation of synaptic plasticity and learning and memory. Neural. Regen. Res. 2022, 17, 705–716.
- Hudson, L.; Liu, J.; Nath, A.; Jones, M.; Raghavan, R.; Narayan, O.; Male, D.; Everall, I. Detection of the human immunodeficiency virus regulatory protein tat in CNS tissues. J. Neurovirol. 2000, 6, 145–155.
- Jones, M.V.; Bell, J.E.; Nath, A. Immunolocalization of HIV envelope gp120 in HIV encephalitis with dementia. AIDS 2000, 14, 2709–2713.
- Wheeler, E.D.; Achim, C.L.; Ayyavoo, V. Immunodetection of human immunodeficiency virus type 1 (HIV-1) Vpr in brain tissue of HIV-1 encephalitic patients. J. Neurovirol. 2006, 12, 200–210.
- Ranki, A.; Nyberg, M.; Ovod, V.; Haltia, M.; Elovaara, I.; Raininko, R.; Haapasalo, H.; Krohn, K. Abundant expression of HIV Nef and Rev proteins in brain astrocytes in vivo is associated with dementia. AIDS 1995, 9, 1001–1008.
- Pushkarsky, T.; Ward, A.; Ivanov, A.; Lin, X.; Sviridov, D.; Nekhai, S.; Bukrinsky, M.I. Abundance of Nef and p-Tau217 in Brains of Individuals Diagnosed with HIV-Associated Neurocognitive Disorders Correlate with Disease Severance. Mol. Neurobiol. 2022, 59, 1088–1097.
- Donoso, M.; D’Amico, D.; Valdebenito, S.; Hernandez, C.A.; Prideaux, B.; Eugenin, E.A. Identification, Quantification, and Characterization of HIV-1 Reservoirs in the Human Brain. Cells 2022, 11, 2379.
- Everall, I.P.; Heaton, R.K.; Marcotte, T.D.; Ellis, R.J.; McCutchan, J.A.; Atkinson, J.H.; Grant, I.; Mallory, M.; Masliah, E. Cortical synaptic density is reduced in mild to moderate human immunodeficiency virus neurocognitive disorder. HNRC Group. HIV Neurobehavioral Research Center. Brain Pathol. 1999, 9, 209–217.
- Tavazzi, E.; Morrison, D.; Sullivan, P.; Morgello, S.; Fischer, T. Brain inflammation is a common feature of HIV-infected patients without HIV encephalitis or productive brain infection. Curr. HIV Res. 2014, 12, 97–110.
- Ginsberg, S.D.; Alldred, M.J.; Gunnam, S.M.; Schiroli, C.; Lee, S.H.; Morgello, S.; Fischer, T. Expression profiling suggests microglial impairment in human immunodeficiency virus neuropathogenesis. Ann. Neurol. 2018, 83, 406–417.