Infertility is a worldwide health issue defined by the World Health Organization (WHO) as the inability to establish a pregnancy after 12 months or more of regular and unprotected sexual intercourse. Male infertility etiology can be related to either congenital or acquired factors. The therapeutical approach to male infertility depends on the underlying causes and includes medical and surgical treatments. In recent studies, the potential role of nerve growth factor (NGF) in male reproductive physiology has been proposed. It has been hypothesized that neurotrophins might be involved in testis morphogenesis and regulation of several aspects of spermatogenesis. Moreover, it has been shown that NGF exerts its role on gonadotropin-releasing hormone (GnRH) neurons through the activation of the PKC/p–ERK1/2/p–CREB cascade, which leads to the activation of hypothalamic cells and the consequent activation of hypothalamus–pituitary–gonadal axis (HPG) with the secretion of GnRH.
- male infertility
- oligozoospermia
- asthenozoospermia
- azoospermia
- sperm cryopreservation
- assisted reproduction
- nerve growth factor
- neurotrophins
1. Introduction
1.1. Male Infertility
1.1.1. Epidemiology
1.1.2. Etiology
1.1.3. Treatments
2. NGF
2.1. Neurotrophins
2.2. NGF Expression in the Reproductive System
3. NGF Specific Functions in the Male Reproductive System
3.1. NGF’s Role in Testis Morphogenesis
3.2. NGF Role in Spermatogenesis
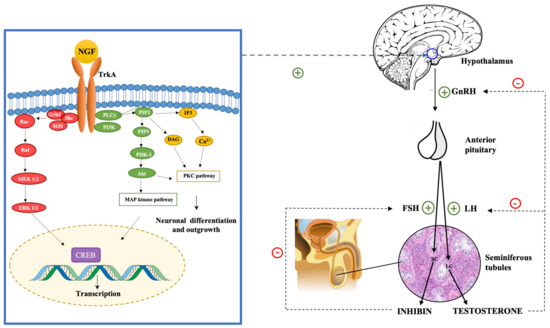
3.3. The Impact of NGF on the Hypothalamus-Pituitary-Gonadal (HPG) Axis
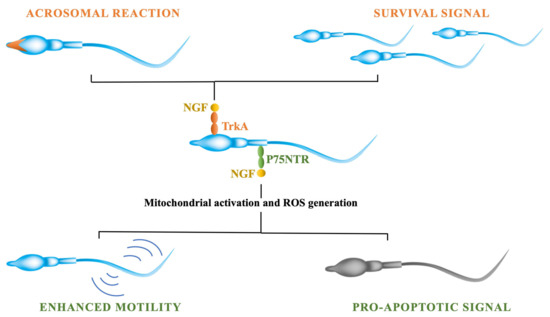
3.4. NGF Effects on Sperm Traits
This entry is adapted from the peer-reviewed paper 10.3390/ijms232113127
References
- Zegers-Hochschild, F.; Adamson, G.D.; Dyer, S.; Racowsky, C.; de Mouzon, J.; Sokol, R.; Rienzi, L.; Sunde, A.; Schmidt, L.; Cooke, I.D.; et al. The International Glossary on Infertility and Fertility Care, 2017. Fertil. Steril. 2017, 108, 393–406.
- Fainberg, J.; Kashanian, J.A. Recent advances in understanding and managing male infertility. F1000Research 2019, 8, 670.
- Vander Borght, M.; Wyns, C. Fertility and infertility: Definition and epidemiology. Clin. Biochem. 2018, 62, 2–10.
- Thoma, M.E.; McLain, A.; Louis, J.F.; King, R.B.; Trumble, A.C.; Sundaram, R.; Louis, G.B. Prevalence of infertility in the United States as estimated by the current duration approach and a traditional constructed approach. Fertil. Steril. 2013, 99, 1324–1331.e1.
- Bak, C.W.; Seok, H.H.; Song, S.H.; Kim, E.S.; Her, Y.S.; Yoon, T.K. Hormonal imbalances and psychological scars left behind in infertile men. J. Androl. 2012, 33, 181–189.
- Wu, A.K.; Elliott, P.; Katz, P.P.; Smith, J.F. Time costs of fertility care: The hidden hardship of building a family. Fertil. Steril. 2013, 99, 2025–2030.
- Dimitriadis, F.; Adonakis, G.; Kaponis, A.; Mamoulakis, C.; Takenaka, A.; Sofikitis, N. Pre-Testicular, Testicular, and Post-Testicular Causes of Male Infertility. In Endocrinology of the Testis and Male Reproduction; Simoni, M., Huhtaniemi, I., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–47.
- Lenzi, A.; Balercia, G.; Bellastella, A.; Colao, A.; Fabbri, A.; Foresta, C.; Galdiero, M.; Gandini, L.; Krausz, C.; Lombardi, G.; et al. Epidemiology; diagnosis, and treatment of male hypogonadotropic hypogonadism. J. Endocrinol. Investig. 2009, 32, 934–938.
- Huhtaniemi, I.; Alevizaki, M. Mutations along the hypothalamic-pituitary-gonadal axis affecting male reproduction. Reprod. Biomed. Online 2007, 15, 622–632.
- Bianco, S.D.C.; Kaiser, U.B. The genetic and molecular basis of idiopathic hypogonadotropic hypogonadism. Nat. Rev. Endocrinol. 2009, 5, 569–576.
- Whitcomb, R.W.; Crowley, W.F. Male hypogonadotropic hypogonadism. Endocrinol. Metab. Clin. N. Am. 1993, 22, 125–143.
- Fraietta, R.; Zylberstejn, D.S.; Esteves, S.C. Hypogonadotropic hypogonadism revisited. Clinics 2013, 68 (Suppl. S1), 81–88.
- Aust, T.R.; Lewis-Jones, D.I. Retrograde ejaculation and male infertility. Hosp. Med. 2004, 65, 361–364.
- Kondoh, N. Ejaculatory dysfunction as a cause of infertility. Reprod. Med. Biol. 2012, 11, 59–64.
- Lotti, F.; Maggi, M. Sexual dysfunction and male infertility. Nat. Rev. Urol. 2018, 15, 287–307.
- Wiser, H.J.; Sandlow, J.; Köhler, T.S. Causes of male infertility. In Male Infertility; Parekattil, S., Agarwal, A., Eds.; Springer: New York, NY, USA, 2012; pp. 3–14.
- Wong, T.W.; Straus, F.H.; Jones, T.M.; Warner, N.E. Pathological aspects of the infertile testis. Urol. Clin. N. Am. 1978, 5, 503–530.
- Cocuzza, M.; Alvarenga, C.; Pagani, R. The epidemiology and etiology of azoospermia. Clinics 2013, 68, 15–26.
- Merino, G.; Carranza-Lira, S.; Murrieta, S.; Rodriguez, L.; Cuevas, E.; Moran, C. Bacterial infection and semen characteristics in infertile men. Syst. Biol. Reprod. Med. 1995, 35, 43–47.
- Diemer, T.; Huwe, P.; Ludwig, M.; Hauck, E.W.; Weidner, W. Urogenital infection and sperm motility. Andrologia 2003, 35, 283–287.
- Krausz, C.; Mills, C.; Rogers, S.; Tan, S.L.; Aitken, R.J. Stimulation of oxidant generation by human sperm suspensions using phorbol esters and formyl peptides: Relationships with motility and fertilization in vitro. Fertil. Steril. 1994, 62, 599–605.
- Restrepo, B.; Cardona Maya, W. Antisperm antibodies and fertility asociation. Actas Urol. Esp. 2013, 37, 571–578.
- Bieth, E.; Hamdi, S.M.; Mieusset, R. Genetics of the congenital absence of the vas deferens. Hum. Genet. 2021, 140, 59–76.
- Sharma, A.; Minhas, S.; Dhillo, W.S.; Jayasena, C.N. Male infertility due to testicular disorders. J. Clin. Endocrinol. Metab. 2021, 106, E442–E459.
- Villumsen, A.L.; Zachau-Christiansen, B. Spontaneous alterations in position of the testes. Arch. Dis. Child. 1966, 41, 198–200.
- Wohlfahrt-Veje, C.; Boisen, K.A.; Boas, M.; Damgaard, I.N.; Kai, C.M.; Schmidt, I.M.; Chellakooty, M.; Suomi, A.-M.; Toppari, J.; Skakkebaek, N.E.; et al. Acquired cryptorchidism is frequent in infancy and childhood. Int. J. Androl. 2009, 32, 423–428.
- Main, K.M.; Skakkebæk, N.E.; Virtanen, H.E.; Toppari, J. Genital anomalies in boys and the environment. Best Pract. Res. Clin. Endocrinol. Metab. 2010, 24, 279–289.
- Kantartzi, P.D.; Goulis, C.D.; Goulis, G.D.; Papadimas, I. Male infertility and varicocele: Myths and reality. Hippokratia 2007, 11, 99–104.
- Ferlin, A.; Arredi, B.; Foresta, C. Genetic causes of male infertility. Reprod. Toxicol. 2006, 22, 133–141.
- Linn, E.; Ghanem, L.; Bhakta, H.; Greer, C.; Avella, M. Genes Regulating Spermatogenesis and Sperm Function Associated With Rare Disorders. Front. Cell Dev. Biol. 2021, 9, 634536.
- Leaver, R.B. Male infertility: An overview of causes and treatment options. Br. J. Nurs. 2016, 25, S35–S40.
- Kamischke, A.; Nieschlag, E. Analysis of medical treatment of male infertility. Hum. Reprod. 1999, 14, 1–23.
- Dabaja, A.A.; Schlegel, P.N. Medical treatment of male infertility. Transl. Androl. Urol. 2014, 3, 9–16.
- Nachtigall, L.B.; Boepple, P.A.; Pralong, F.P.; Crowley, W.F. Adult-Onset Idiopathic Hypogonadotropic Hypogonadism—A Treatable Form of Male Infertility. N. Engl. J. Med. 1997, 336, 410–415.
- Conn, P.M.; Hansen, J.R. Gonadotropin-releasing hormone and its analogs. Iowa Med. 1986, 76, 372–377.
- Zitzmann, M.; Nieschlag, E. Hormone substitution in male hypogonadism. Mol. Cell. Endocrinol. 2000, 161, 73–88.
- Blumenfeld, Z.; Frisch, L.; Conn, P.M. Gonadotropin-releasing hormone (GnRH) antibodies formation in hypogonadotropic azoospermic men treated with pulsatile GnRH-Diagnosis and possible alternative treatment. Fertil. Steril. 1988, 50, 622–629.
- Wei, C.; Long, G.; Zhang, Y.; Wang, T.; Wang, S.; Liu, J.; Ma, D.; Liu, X. Spermatogenesis of male patients with congenital hypogonadotropic hypogonadism receiving pulsatile gonadotropin-releasing hormone therapy versus gonadotropin therapy: A systematic review and meta-analysis. World J. Mens Health 2020, 38, 654–665.
- Liu, L.; Banks, S.M.; Barnes, K.M.; Sherins, R.J. Two-year comparison of testicular responses to pulsatile gonadotropin-releasing hormone and exogenous gonadotropins from the inception of therapy in men with isolated hypogonadotropic hypogonadism. J. Clin. Endocrinol. Metab. 1988, 67, 1140–1145.
- Madhukar, D.; Rajender, S. Hormonal treatment of male infertility: Promise and pitfalls. J. Androl. 2009, 30, 95–112.
- Burgués, S.; Calderón, M.D. Subcutaneous self-administration of highly purified follicle stimulating hormone and human chorionic gonadotrophin for the treatment of male hypogonadotrophic hypogonadism. Hum. Reprod. 1997, 12, 980–986.
- Dabbous, Z.; Atkin, S.L. Hyperprolactinaemia in male infertility: Clinical case scenarios. Arab J. Urol. 2018, 16, 44–52.
- Cannarella, R.; Condorelli, R.A.; Mongioì, L.M.; Barbagallo, F.; Calogero, A.E.; La Vignera, S. Effects of the selective estrogen receptor modulators for the treatment of male infertility: A systematic review and meta-analysis. Expert Opin. Pharmacother. 2019, 20, 1517–1525.
- Goldstein, S.R.; Siddhanti, S.; Ciaccia, A.V.; Plouffe, L. A pharmacological review of selective oestrogen receptor modulators. Hum. Reprod. Update 2000, 6, 212–224.
- Velasquez, M.; Tanrikut, C. Surgical management of male infertility: An update. Transl. Androl. Urol. 2014, 3, 64–76.
- Schiff, J.D.; Ramírez, M.L.; Bar-Chama, N. Medical and Surgical Management Male Infertility. Endocrinol. Metab. Clin. N. Am. 2007, 36, 313–331.
- Tournaye, H. Male factor infertility and ART. Asian J. Androl. 2012, 14, 103–108.
- Duran, H.E.; Morshedi, M.; Kruger, T.; Oehninger, S. Intrauterine insemination: A systematic review on determinants of success. Hum. Reprod. Update 2002, 8, 373–384.
- Huang, J.Y.J.; Rosenwaks, Z. Assisted reproductive techniques. Methods Mol. Biol. 2014, 1154, 171–231.
- Aboulghar, M.; Baird, D.T.; Collins, J.; Evers, J.L.H.; Fauser, B.C.J.M.; Lambalk, C.B.; Somigliana, E.; Sunde, A.; Tarlatzis, B.; Crosignani, P.G.; et al. Intrauterine insemination. Hum. Reprod. Update 2009, 15, 265–277.
- Keck, C.; Gerber-Schafer, C.; Wilhelm, C.; Vogelgesang, D.; Breckwoldt, M. Intrauterine insemination for treatment of male infertility. Int. J. Androl. Suppl. 1997, 20, 55–64.
- Hasler, J.F.; Barfield, J.P. Vitro Fertilization. In Bovine Reproduction; StatPearls Publishing LLC.: Treasure Island, FL, USA, 2021; pp. 1124–1141.
- Palermo, G.D.; Neri, Q.V.; Hariprashad, J.J.; Davis, O.K.; Veeck, L.L.; Rosenwaks, Z. ICSI and its outcome. Semin. Reprod. Med. 2000, 18, 161–169.
- O’Neill, C.L.; Chow, S.; Rosenwaks, Z.; Palermo, G.D. Development of ICSI. Reproduction 2018, 156, F51–F58.
- Justice, T.; Christensen, G. Sperm Cryopreservation Methods. Methods Mol. Biol. 2013, 927, 209–215.
- Skaper, S.D. Neurotrophic Factors: An Overview. Methods Mol. Biol. 2018, 1727, 1–17.
- Bothwell, M. Keeping track of neurotrophin receptors. Cell 1991, 65, 915–918.
- Chao, M.V. Neurotrophins and their receptors: A convergence point for many signalling pathways. Nat. Rev. Neurosci. 2003, 4, 299–309.
- Casaccia-Bonnefil, P.; Carter, B.D.; Dobrowsky, R.T.; Chao, M.V. Death of oligodendrocytes mediated by the interaction of nerve growth factor with its receptor p75. Nature 1996, 383, 716–719.
- Friedman, W.J.; Greene, L.A. Neurotrophin signaling via Trks and p75. Exp. Cell Res. 1999, 253, 131–142.
- Pearson, G.; Robinson, F.; Beers Gibson, T.; Xu, B.E.; Karandikar, M.; Berman, K.; Cobb, M.H. Mitogen-Activated Protein (MAP) Kinase Pathways: Regulation and Physiological Functions. Endocr. Rev. 2001, 22, 153–183.
- Yuan, X.-B.; Jin, M.; Xu, X.; Song, Y.-Q.; Wu, C.-P.; Poo, M.-M.; Duan, S. Signalling and crosstalk of Rho GTPases in mediating axon guidance. Nat. Cell Biol. 2003, 5, 38–45.
- Rose, C.R.; Blum, R.; Pichler, B.; Lepier, A.; Kafitz, K.W.; Konnerth, A. Truncated TrkB-T1 mediates neurotrophin-evoked calcium signalling in glia cells. Nature 2003, 426, 74–78.
- Toledo-Aral, J.J.; Brehm, P.; Halegoua, S.; Mandel, G. A single pulse of nerve growth factor triggers long-term neuronal excitability through sodium channel gene induction. Neuron 1995, 14, 607–611.
- Minichiello, L.; Calella, A.M.; Medina, D.L.; Bonhoeffer, T.; Klein, R.; Korte, M. Mechanism of TrkB-mediated hippocampal long-term potentiation. Neuron 2002, 36, 121–137.
- Nahum, L.H. the Nerve Growth Factor (Ngf). Conn. Med. 1964, 28, 508–512.
- Fiore, M.; Amendola, T.; Triaca, V.; Tirassa, P.; Alleva, E.; Aloe, L. Agonistic encounters in aged male mouse potentiate the expression of endogenous brain NGF and BDNF: Possible implication for brain progenitor cells’ activation. Eur. J. Neurosci. 2003, 17, 1455–1464.
- Ciafrè, S.; Ferraguti, G.; Tirassa, P.; Iannitelli, A.; Ralli, M.; Greco, A.; Chaldakov, G.N.; Rosso, P.; Fico, E.; Messina, M.P.; et al. Nerve growth factor in the psychiatric brain. Riv. Psichiatr. 2020, 55, 4–15.
- Wood, S.J.; Pritchard, J.; Sofroniew, M.V. Re-expression of Nerve Growth Factor Receptor after Axonal Injury Recapitulates a Developmental Event in Motor Neurons: Differential Regulation when Regeneration is Allowed or Prevented. Eur. J. Neurosci. 1990, 2, 650–657.
- Verge, V.M.K.; Richardson, P.M.; Benoit, R.; Riopelle, R.J. Histochemical characterization of sensory neurons with high-affinity receptors for nerve growth factor. J. Neurocytol. 1989, 18, 583–591.
- Ruit, K.G.; Osborne, P.A.; Schmidt, R.E.; Johnson, E.M.; Snider, W.D. Nerve growth factor regulates sympathetic ganglion cell morphology and survival in the adult mouse. J. Neurosci. 1990, 10, 2412–2419.
- Heumann, R.; Lindholm, D.; Bandtlow, C.; Meyer, M.; Radeke, M.J.; Misko, T.P.; Shooter, E.; Thoenen, H. Differential regulation of mRNA encoding nerve growth factor and its receptor in rat sciatic nerve during development, degeneration, and regeneration: Role of macrophages. Proc. Natl. Acad. Sci. USA 1987, 84, 8735–8739.
- Chaldakov, G.N.; Stankulov, I.S.; Fiore, M.; Ghenev, P.I.; Aloe, L. Nerve growth factor levels and mast cell distribution in human coronary atherosclerosis. Atherosclerosis 2001, 159, 57–66.
- Aloe, L.; Alleva, E.; Fiore, M. Stress and nerve growth factor: Findings in animal models and humans. Pharmacol. Biochem. Behav. 2002, 73, 159–166.
- Schulte-Herbruggen, O.; Braun, A.; Rochlitzer, S.; Jockers-Scherubl, M.C.; Hellweg, R. Neurotrophic factors—A tool for therapeutic strategies in neurological, neuropsychiatric and neuroimmunological diseases? Curr. Med. Chem. 2007, 14, 2318–2329.
- Bruscolini, A.; Sacchetti, M.; La Cava, M.; Nebbioso, M.; Iannitelli, A.; Quartini, A.; Lambiase, A.; Ralli, M.; de Virgilio, A.; Greco, A. Quality of life and neuropsychiatric disorders in patients with Graves’ Orbitopathy: Current concepts. Autoimmun. Rev. 2018, 17, 639–643.
- Chaldakov, G.N.; Fiore, M.; Tonchev, A.B.; Aloe, L. Neuroadipology: A novel component of neuroendocrinology. Cell Biol. Int. 2010, 34, 1051–1053.
- Tore, F.; Tonchev, A.; Fiore, M.; Tuncel, N.; Atanassova, P.; Aloe, L.; Chaldakov, G. From Adipose Tissue Protein Secretion to Adipopharmacology of Disease. Immunol. Endocr. Metab. Agents Med. Chem. 2007, 7, 149–155.
- Carito, V.; Ceccanti, M.; Tarani, L.; Ferraguti, G.; Chaldakov, G.N.; Fiore, M. Neurotrophins’ Modulation by Olive Polyphenols. Curr. Med. Chem. 2016, 23, 3189–3197.
- Petrella, C.; Di Certo, M.G.; Gabanella, F.; Barbato, C.; Ceci, F.M.; Greco, A.; Ralli, M.; Polimeni, A.; Angeloni, A.; Severini, C.; et al. Mediterranean Diet, Brain and Muscle: Olive Polyphenols and Resveratrol Protection in Neurodegenerative and Neuromuscular Disorders. Curr. Med. Chem. 2021, 28, 7595–7613.
- Ceci, F.M.; Ferraguti, G.; Petrella, C.; Greco, A.; Tirassa, P.; Iannitelli, A.; Ralli, M.; Vitali, M.; Ceccanti, M.; Chaldakov, G.N.; et al. Nerve Growth Factor, Stress and Diseases. Curr. Med. Chem. 2020, 28, 2943–2959.
- Rosso, P.; Iannitelli, A.; Pacitti, F.; Quartini, A.; Fico, E.; Fiore, M.; Greco, A.; Ralli, M.; Tirassa, P. Vagus nerve stimulation and Neurotrophins: A biological psychiatric perspective. Neurosci. Biobehav. Rev. 2020, 113, 338–353.
- Carito, V.; Ceccanti, M.; Ferraguti, G.; Coccurello, R.; Ciafrè, S.; Tirassa, P.; Fiore, M. NGF and BDNF Alterations by Prenatal Alcohol Exposure. Curr. Neuropharmacol. 2019, 17, 308–317.
- Ciafrè, S.; Ferraguti, G.; Greco, A.; Polimeni, A.; Ralli, M.; Ceci, F.M.; Ceccanti, M.; Fiore, M. Alcohol as an early life stressor: Epigenetics, metabolic, neuroendocrine and neurobehavioral implications. Neurosci. Biobehav. Rev. 2020, 118, 654–668.
- Ceci, F.M.; Ferraguti, G.; Petrella, C.; Greco, A.; Ralli, M.; Iannitelli, A.; Carito, V.; Tirassa, P.; Chaldakov, G.N.; Messina, M.P.; et al. Nerve Growth Factor in Alcohol Use Disorders. Curr. Neuropharmacol. 2020, 19, 45–60.
- Fiore, M.; Triaca, V.; Amendola, T.; Tirassa, P.; Aloe, L. Brain NGF and EGF administration improves passive avoidance response and stimulates brain precursor cells in aged male mice. Physiol. Behav. 2002, 77, 437–443.
- Chaldakov, G.; Fiore, M.; Tonchev, A.; Dimitrov, D.; Pancheva, R.; Rancic, G.; Aloe, L. Homo obesus: A Metabotrophin-Deficient Species. Pharmacol. Nutr. Insight Curr. Pharm. Des. 2007, 13, 2176–2179.
- Miranda, M.; Morici, J.F.; Zanoni, M.B.; Bekinschtein, P. Brain-Derived Neurotrophic Factor: A Key Molecule for Memory in the Healthy and the Pathological Brain. Front. Cell. Neurosci. 2019, 13, 363.
- Otten, U.; Ehrhard, P.; Peck, R. Nerve growth factor induces growth and differentiation of human B lymphocytes. Proc. Natl. Acad. Sci. USA 1989, 86, 10059–10063.
- Torcia, M.; Bracci-Laudiero, L.; Lucibello, M.; Nencioni, L.; Labardi, D.; Rubartelli, A.; Cozzolino, F.; Aloe, L.; Garaci, E. Nerve growth factor is an autocrine survival factor for memory B lymphocytes. Cell 1996, 85, 345–356.
- Kannan, Y.; Usami, K.; Okada, M.; Shimizu, S.; Matsuda, H. Nerve growth factor suppresses apoptosis of murine neutrophils. Biochem. Biophys. Res. Commun. 1992, 186, 1050–1056.
- Horigome, K.; Bullock, E.D.; Johnson, E.M. Effects of nerve growth factor on rat peritoneal mast cells. Survival promotion and immediate-early gene induction. J. Biol. Chem. 1994, 269, 2695–2702.
- Shaoxia, P.U.; Changwei, Q.U.; Zhi, L.I.; Yansen, L.I.; Chunmei, L.I. Expression of nerve growth factor (NGF) and its receptors TrkA and p75 in the reproductive organs of laying hens. Rev. Bras. Cienc. Avic. 2016, 18, 187–192.
- Perrard, M.-H.; Vigier, M.; Damestoy, A.; Chapat, C.; Silandre, D.; Rudkin, B.B.; Durand, P. Β-Nerve Growth Factor Participates in an Auto/Paracrine Pathway of Regulation of the Meiotic Differentiation of Rat Spermatocytes. J. Cell. Physiol. 2007, 210, 51–62.
- Artico, M.; Bronzetti, E.; Saso, L.; Felici, L.M.; D’Ambrosio, A.; Forte, F.; Grande, C.; Ortolani, F. Immunohistochemical profile of some neurotransmitters and neurotrophins in the seminiferous tubules of rats treated by lonidamine. Eur. J. Histochem. 2007, 51, 19–24.
- Adams, G.P.; Ratto, M.H. Ovulation-inducing factor in seminal plasma: A review. Anim. Reprod. Sci. 2013, 136, 148–156.
- Hou, Y.; Jia, L.; Zhang, Y.; Ji, W.; Li, H. Activation of the NGF/TrkA signaling pathway attenuates diabetic erectile dysfunction. Oncotarget 2017, 8, 105692–105702.
- Spinnler, K.; Fröhlich, T.; Arnold, G.J.; Kunz, L.; Mayerhofer, A. Human tryptase cleaves pro-nerve growth factor (Pro-NGF): Hints of local, mast cell-dependent regulation of NGF/PRO-NGF action. J. Biol. Chem. 2011, 286, 31707–31713.
- Guo, J.; Zhu, P.; Wu, C.; Yu, L.; Zhao, S.; Gu, X. In silico analysis indicates a similar gene expression pattern between human brain and testis. Cytogenet. Genome Res. 2003, 103, 58–62.
- Graves, J.A.M. Review: Sex Chromosome Evolution and the Expression of Sex-Specific Genes in the Placenta. Placenta 2010, 31, S27–S32.
- Ramírez-Reveco, A.; Villarroel-Espíndola, F.; Rodríguez-Gil, J.E.; Concha, I.I. Neuronal signaling repertoire in the mammalian sperm functionality. Biol. Reprod. 2017, 96, 505–524.
- Ayer-LeLievre, C.; Olson, L.; Ebendal, T.; Hallbook, F.; Persson, H. Nerve growth factor mRNA and protein in the testis and epididymis of mouse and rat. Proc. Natl. Acad. Sci. USA 1988, 85, 2628–2632.
- Harper, G.P.; Barde, Y.A.; Burnstock, G.; Carstairs, J.R.; Dennison, M.E.; Suda, K.; Vernon, C.A. Guinea pig prostate is a rich source of nerve growth factor. Nature 1979, 279, 160–162.
- Harper, G.P.; Thoenen, H. The Distribution of Nerve Growth Factor in the Male Sex Organs of Mammals. J. Neurochem. 1980, 34, 893–903.
- Jin, W.Z.; Tanaka, A.; Watanabe, G.; Matsuda, H.; Taya, K. Effect of NGF on the motility and acrosome reaction of golden hamster spermatozoa in vitro. J. Reprod. Dev. 2010, 56, 437–443.
- Jin, W.; Arai, K.Y.; Shimizu, K.; Kojima, C.; Itoh, M.; Watanabe, G.; Taya, K. Cellular localization of NGF and its receptors trkA and p75LNGFR in male reproductive organs of the Japanese monkey, Macaca fuscata fuscata. Endocrine 2006, 29, 155–160.
- Levanti, M.B.; Germanà, A.; de Carlos, F.; Ciriaco, E.; Vega, J.A.; Germanà, G. Effects of increased nerve growth factor plasma levels on the expression of TrkA and p75NTR in rat testicles. J. Anat. 2006, 208, 373–379.
- Kumar, S.; Sharma, V.K.; Singh, S.; Hariprasad, G.R.; Mal, G.; Srinivasan, A.; Yadav, S. Proteomic identification of camel seminal plasma: Purification of β-nerve growth factor. Anim. Reprod. Sci. 2013, 136, 289–295.
- Druart, X.; Rickard, J.; Mactier, S.; Kohnke, P.; Kershaw-Young, C.; Bathgate, R.; Gibb, Z.; Crossett, B.; Tsikis, G.; Labas, V.; et al. Proteomic characterization and cross species comparison of mammalian seminal plasma. J. Proteomics 2013, 91, 13–22.
- Harper, G.P.; Glanville, R.W.; Thoenen, H. The purification of nerve growth factor from bovine seminal plasma. Biochemical characterization and partial amino acid sequence. J. Biol. Chem. 1982, 257, 8541–8548.
- Li, C.; Zheng, L.; Wang, C.; Zhou, X. Absence of nerve growth factor and comparison of tyrosine kinase receptor A levels in mature spermatozoa from oligoasthenozoospermic, asthenozoospermic and fertile men. Clin. Chim. Acta 2010, 411, 1482–1486.
- Adams, G.P.; Ratto, M.H.; Silva, M.E.; Carrasco, R.A. Ovulation-inducing factor (OIF/NGF) in seminal plasma: A review and update. Reprod. Domest. Anim. 2016, 51, 4–17.
- Seidl, K.; Buchberger, A.; Erck, C. Expression of nerve growth factor and neurotrophin receptors in testicular cells suggest novel roles for neurotrophins outside the nervous system. Reprod. Fertil. Dev. 1996, 8, 1075–1087.
- Brill, G.; Kahane, N.; Carmeli, C.; Von Schack, D.; Barde, Y.A.; Kalcheim, C. Epithelial-mesenchymal conversion of dermatome progenitors requires neural tube-derived signals: Characterization of the role of Neurotrophin-3. Development 1995, 121, 2583–2594.
- Dissen, G.A.; Newman Hirshfield, A.; Malamed, S.; Ojeda, S.R. Expression of neurotrophins and their receptors in the mammalian ovary is developmentally regulated: Changes at the time of folliculogenesis. Endocrinology 1995, 136, 4681–4692.
- Mitsiadis, T.A.; Luukko, K. Neurotrophins in odontogenesis. Int. J. Dev. Biol. 1995, 39, 195–202.
- Ojeda, S.R.; Dissen, G.A.; Junier, M.P. Neurotrophic factors and female sexual development. Front. Neuroendocrinol. 1992, 13, 120–162.
- Onoda, M.; Pflug, B.; Djakiew, D. Germ cell mitogenic activity is associated with nerve growth factor-like protein(s). J. Cell. Physiol. 1991, 149, 536–543.
- Persson, H.; Lievre, C.A.-L.; Söder, O.; Villar, M.J.; Metsis, M.; Olson, L.; Ritzen, M.; Hökfelt, T. Expression of β-nerve growth factor receptor mRNA in Sertoli cells downregulated by testosterone. Science 1990, 247, 704–707.
- Lonnerberg, P.; Soder, O.; Parvinen, M.; Ritzen, E.M.; Persson, H. β-Nerve growth factor influences the expression of androgen-binding protein messenger ribonucleic acid in the rat testis. Biol. Reprod. 1992, 47, 381–388.
- Wheeler, E.F.; Bothwell, M. Spatiotemporal patterns of expression of NGF and the low-affinity NGF receptor in rat embryos suggest functional roles in tissue morphogenesis and myogenesis. J. Neurosci. 1992, 12, 930–945.
- Russo, M.A.; Odorisio, T.; Fradeani, A.; Rienzi, L.; De Felici, M.; Cattaneo, A.; Siracusa, G. Low-affinity nerve growth factor receptor is expressed during testicular morphogenesis and in germ cells at specific stages of spermatogenesis. Mol. Reprod. Dev. 1994, 37, 157–166.
- Levine, E.; Cupp, A.S.; Skinner, M.K. Role of neurotropins in rat embryonic testis morphogenesis (Cord formation). Biol. Reprod. 2000, 62, 132–142.
- Cupp, A.S.; Kim, G.H.; Skinner, M.K. Expression and action of neurotropin-3 and nerve growth factor in embryonic and early postnatal rat testis development. Biol. Reprod. 2000, 63, 1617–1628.
- Cupp, A.S.; Tessarollo, L.; Skinner, M.K. Testis developmental phenotypes in neurotropin receptor trkA and trkC null mutations: Role in formation of seminiferous cords and germ cell survival. Biol. Reprod. 2002, 66, 1838–1845.
- Neto, F.T.L.; Bach, P.V.; Najari, B.B.; Li, P.S.; Goldstein, M. Spermatogenesis in humans and its affecting factors. Semin. Cell Dev. Biol. 2016, 59, 10–26.
- Nishimura, H.; L’Hernault, S.W. Spermatogenesis. Curr Biol. 2017, 27, R988–R994.
- De Kretser, D.M.; Loveland, K.L.; Meinhardt, A.; Simorangkir, D.; Wreford, N. Spermatogenesis. Hum. Reprod. 1998, 13, 1–8.
- Larose, H.; Kent, T.; Ma, Q.; Shami, A.N.; Harerimana, N.; Li, J.Z.; Hammoud, S.S.; Handel, M.A. Regulation of meiotic progression by Sertoli-cell androgen signaling. Mol. Biol. Cell 2020, 31, 2841–2862.
- Hess, R.A.; De Franca, L.R. Spermatogenesis and cycle of the seminiferous epithelium. Adv. Exp. Med. Biol. 2008, 636, 1–15.
- Holdcraft, R.W.; Braun, R.E. Hormonal regulation of spermatogenesis. Int. J. Androl. 2004, 27, 335–342.
- Rossi, P.; Dolci, S. Paracrine mechanisms involved in the control of early stages of mammalian spermatogenesis. Front. Endocrinol. 2013, 4, 181.
- Sofikitis, N.; Giotitsas, N.; Tsounapi, P.; Baltogiannis, D.; Giannakis, D.; Pardalidis, N. Hormonal regulation of spermatogenesis and spermiogenesis. J. Steroid Biochem. Mol. Biol. 2008, 109, 323–330.
- Griswold, M.D. 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells. Biol. Reprod. 2018, 99, 87–100.
- Schlatt, S.; Meinhardt, A.; Nieschlag, E. Paracrine regulation of cellular interactions in the testis: Factors in search of a function. Eur. J. Endocrinol. 1997, 137, 107–117.
- Iliadou, P.K.; Tsametis, C.; Kaprara, A.; Papadimas, I.; Goulis, D.G. The sertoli cell: Novel clinical potentiality. Hormones 2015, 14, 504–514.
- Galdieri, M.; Monaco, L.; Stefanini, M. Secretion of Androgen Binding Protein by Sertoli Cells Is Influenced by Contact with Germ Cells. J. Androl. 1984, 5, 409–415.
- Le Magueresse, B.; Jégou, B. In vitro effects of germ cells on the secretory activity of sertoli cells recovered from rats of different ages. Endocrinology 1988, 122, 1672–1680.
- Onoda, M.; Djakiew, D. A 29,000 Mr protein derived from round spermatids regulates Sertoli cell secretion. Mol. Cell. Endocrinol. 1993, 93, 53–61.
- Haugen, T.B.; Landmark, B.F.; Josefsen, G.M.; Hansson, V.; Högset, A. The mature form of interleukin-1α is constitutively expressed in immature male germ cells from rat. Mol. Cell. Endocrinol. 1994, 105, R19–R23.
- Onoda, M.; Djakiew, D. Pachytene spermatocyte protein(s) stimulate sertoli cells grown in bicameral chambers: Dose-dependent secretion of ceruloplasmin, sulfated glycoprotein-1, sulfated glycoprotein-2, and transferrin. Vitr. Cell. Dev. Biol.-Anim. 1991, 27, 215–222.
- Pineau, C.; Sharpe, R.M.; Saunders, P.T.K.; Gérard, N.; Jégou, B. Regulation of Sertoli cell inhibin production and of inhibin α-subunit mRNA levels by specific germ cell types. Mol. Cell. Endocrinol. 1990, 72, 13–22.
- Onoda, M.; Djakiew, D. Modulation of Sertoli cell secretory function by rat round spermatid protein(s). Mol. Cell. Endocrinol. 1990, 73, 35–44.
- Le Magueresse, B.; Jegou, B. Possible involvement of germ cells in the regulation of oestradiol-17,β and ABP secretion by immature rat sertoli cells (in vitro studies). Biochem. Biophys. Res. Commun. 1986, 141, 861–869.
- Vigier, M.; Weiss, M.; Perrard, M.H.; Godet, M.; Durand, P. The effects of FSH and of testosterone on the completion of meiosis and the very early steps of spermiogenesis of the rat: An in vitro study. J. Mol. Endocrinol. 2004, 33, 729–742.
- Hakovirta, H.; Kaipia, A.; Söder, O.; Parvinen, M. Effects of activin-A, inhibin-A, and transforming growth factor-β1 on stage-specific deoxyribonucleic acid synthesis during rat seminiferous epithelial cycle. Endocrinology 1993, 133, 1664–1668.
- Olson, L.; Ayer-LeLievre, C.; Ebendal, T.; Seiger, Å. Nerve growth factor-like immunoreactivities in rodent salivary glands and testis. Cell Tissue Res. 1987, 248, 275–286.
- MacGrogan, D.; Desprès, G.; Romand, R.; Dicou, E. Expression of the β-nerve growth factor gene in male sex organs of the mouse, rat, and guinea pig. J. Neurosci. Res. 1991, 28, 567–573.
- Seidl, K.; Holstein, A.F. Organ culture of human seminiferous tubules: A useful tool to study the role of nerve growth factor in the testis. Cell Tissue Res. 1990, 261, 539–547.
- Parvinen, M.; Pelto-Huikko, M.; Soder, O.; Schultz, R.; Kaipia, A.; Mali, P.; Toppari, J.; Hakovirta, H.; Lönnerberg, P.; Ritzén, E.M. Expression of β-nerve growth factor and its receptor in rat seminiferous epithelium: Specific function at the onset of meiosis. J. Cell Biol. 1992, 117, 629–641.
- Djakiew, D.; Pflug, B.; Dionne, C.; Onoda, M. Postnatal expression of nerve growth factor receptors in the rat testis. Biol. Reprod. 1994, 51, 214–221.
- MacGrogan, D.; Saint-André, J.-P.; Dicou, E. Expression of Nerve Growth Factor and Nerve Growth Factor Receptor Genes in Human Tissues and in Prostatic Adenocarcinoma Cell Lines. J. Neurochem. 1992, 59, 1381–1391.
- Robinson, L.L.L.; Townsend, J.; Anderson, R.A. The human fetal testis is a site of expression of neurotrophins and their receptors: Regulation of the germ cell and peritubular cell population. J. Clin. Endocrinol. Metab. 2003, 88, 3943–3951.
- Plant, T.M. The hypothalamo-pituitary-gonadal axis. J. Endocrinol. 2015, 226, T41–T54.
- Jin, J.M.; Yang, W.X. Molecular regulation of hypothalamus-pituitary-gonads axis in males. Gene 2014, 551, 15–25.
- Stamatiades, G.A.; Kaiser, U.B. Gonadotropin regulation by pulsatile GnRH: Signaling and gene expression. Mol. Cell. Endocrinol. 2018, 463, 131–141.
- Zirkin, B.R.; Papadopoulos, V. Leydig cells: Formation, function, and regulation. Biol. Reprod. 2018, 99, 101–111.
- Oduwole, O.O.; Peltoketo, H.; Huhtaniemi, I.T. Role of follicle-stimulating hormone in spermatogenesis. Front. Endocrinol. 2018, 9, 763.
- Ramaswamy, S.; Weinbauer, G.F. Endocrine control of spermatogenesis: Role of FSH and LH/testosterone. Spermatogenesis 2014, 4, e996025.
- Tilbrook, A.J.; Clarke, I.J. Negative feedback regulation of the secretion and actions of gonadotropin-releasing hormone in males. Biol. Reprod. 2001, 64, 735–742.
- Corradi, P.F.; Corradi, R.B.; Greene, L.W. Physiology of the Hypothalamic Pituitary Gonadal Axis in the Male. Urol. Clin. N. Am. 2016, 43, 151–162.
- Luisi, S.; Florio, P.; Reis, F.M.; Petraglia, F. Inhibins in female and male reproductive physiology: Role in gametogenesis, conception, implantation and early pregnancy. Hum. Reprod. Update 2005, 11, 123–135.
- Scaccianoce, S.; Cigliana, G.; Nicolai, R.; Muscolo, L.A.; Porcu, A.; Navarra, D.; Perez-Polo, R.; Angelucci, L. Hypothalamic involvement in the activation of the pituitary-adrenocortical axis by nerve growth factor. Neuroendocrinology 1993, 58, 202–209.
- Kumar, A.; Kumar, P.; Pareek, V.; Faiq, M.A.; Narayan, R.K.; Raza, K.; Prasoon, P.; Sharma, V.K. Neurotrophin mediated HPA axis dysregulation in stress induced genesis of psychiatric disorders: Orchestration by epigenetic modifications. J. Chem. Neuroanat. 2019, 102, 101688.
- Luo, J.; Yang, Y.; Zhang, T.; Su, Z.; Yu, D.; Lin, Q.; Chen, H.; Zhang, Q.; Xiang, Q.; Xue, W.; et al. Nasal delivery of nerve growth factor rescue hypogonadism by up-regulating GnRH and testosterone in aging male mice. EBioMedicine 2018, 35, 295–306.
- Marlin, M.C.; Li, G. Biogenesis and Function of the NGF/TrkA Signaling Endosome. Int. Rev. Cell Mol. Biol. 2015, 314, 239–257.
- Higa-Nakamine, S.; Maeda, N.; Toku, S.; Yamamoto, H. Involvement of protein kinase D1 in signal transduction from the protein kinase C pathway to the tyrosine kinase pathway in response to gonadotropin-releasing hormone. J. Biol. Chem. 2015, 290, 25974–25985.
- Sasson, R.; Dearth, R.K.; White, R.S.; Chappell, P.E.; Mellon, P.L. Orexin A induces GnRH gene expression and secretion from GT1-7 hypothalamic GnRH neurons. Neuroendocrinology 2007, 84, 353–363.
- Riccio, A.; Ahn, S.; Davenport, C.M.; Blendy, J.A.; Ginty, D.D. Mediation by a CREB family transcription factor of NGF-dependent survival of sympathetic neurons. Science 1999, 286, 2358–2361.
- Liu, Y.Z.; Chrivia, J.C.; Latchman, D.S. Nerve growth factor up-regulates the transcriptional activity of CBP through activation of the p42/p44(MAPK) cascade. J. Biol. Chem. 1998, 273, 32400–32407.
- Wortzel, I.; Seger, R. The ERK cascade: Distinct functions within various subcellular organelles. Genes Cancer 2011, 2, 195–209.
- Sanchez-Rodriguez, A.; Abad, P.; Arias-Alvarez, M.; Rebollar, P.G.; Bautista, J.M.; Lorenzo, P.L.; García-García, R.M. Recombinant rabbit beta nerve growth factor production and its biological effects on sperm and ovulation in rabbits. PLoS ONE 2019, 14, e0219780.
- Li, C.; Zhou, X. The potential roles of neurotrophins in male reproduction. Reproduction 2013, 145, R89–R95.
- Li, C.; Sun, Y.; Yi, K.; Ma, Y.; Zhang, W.; Zhou, X. Detection of nerve growth factor (NGF) and its specific receptor (TrkA) in ejaculated bovine sperm, and the effects of NGF on sperm function. Theriogenology 2010, 74, 1615–1622.
- Shi, C.-G.; Lin, K.; Xu, X.-B.; Zhang, S.-C.; Wang, N.; Fan, M. Evidence for the involvement of NGF in human sperm motility. J. Biomed. Sci. Eng. 2012, 5, 534–541.
- Lin, K.; Ding, X.-F.; Shi, C.-G.; Zeng, D.; QuZong, S.; Liu, S.-H.; Wu, Y.; LuoBu, G.; Fan, M.; Zhao, Y.-Q. Nerve growth factor promotes human sperm motility in vitro by increasing the movement distance and the number of A grade spermatozoa. Andrologia 2015, 47, 1041–1046.
- Bezerra, M.; Arruda-Alencar, J.; Martins, J.; Viana, A.; Neto, A.V.; Rêgo, J.; de Oliveira, R.V.; Lobo, M.; Moreira, A.; Moreira, R.; et al. Major seminal plasma proteome of rabbits and associations with sperm quality. Theriogenology 2019, 128, 156–166.