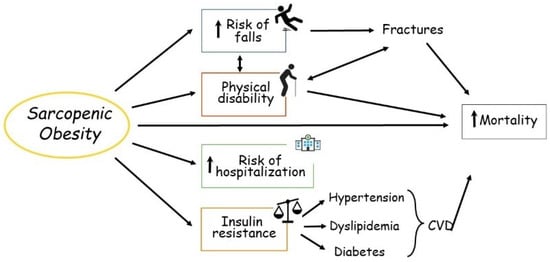
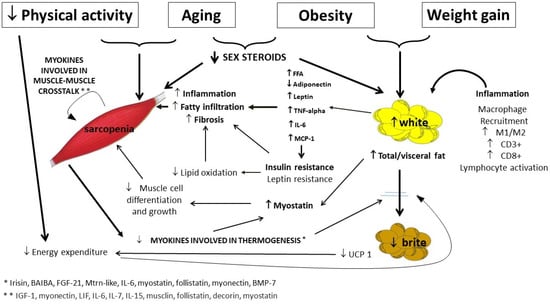
Gain in adipose tissue (WAT) and especially dysfunctional WAT may represent an independent determinant for the development of loss and dysfunction of muscle mass. In addition, a decline in muscle mass may facilitate WAT accumulation. SO is more frequently present in older adults, particularly because of the changes observed in body composition (i.e., changes in muscle mass and AT quantity and quality), which in general accompany the aging process.
2. Adipose Tissue as Endocrine Organ
The concept that WAT, apart from serving as an energy storage and mechanical protection, acts as an endocrine organ, representing a source of countless adipokines, proteins, metabolites, lipid molecules, non-coding RNAs and extracellular vesicles (Evs), as involved in tissue crosstalk, has been widely accepted in recent years [
6]. Besides adipocytes, WAT contains stromovascular cells and immune cells acting as an integrated unit, all of which contribute to its endocrine activity [
8]. Several adipocyte-derived proteins with endocrine function have been detected since the discovery of leptin many years ago [
6].
WAT, distributed in different depots in the body, expresses specific features in relation to cellular composition and functions [
6]. Furthermore, the anatomical location is relevant when considering that hormones produced by the visceral adipose tissue (VAT) are released into the portal system and go directly to the liver, while the systemic circulation receives the molecules produced by the subcutaneous adipose tissue (SAT) [
8,
9].
Moreover, WAT is a dynamic organ that modifies in response to changes in nutritional state, through modifications of its metabolism and cellularity, and subsequent shifts in adipokine secretion. In the lean state, SAT is characterized by smaller cell size and by an increased number of adipocytes, which have a different pattern of adipokine secretion compared with VAT. Weight gain causes adipocyte hypertrophy, with differences across sex, age and fat depot. Weight gain in women, at least until menopause, characterized by the enhanced accumulation of SAT, is associated with a low risk of type 2 diabetes and cardiovascular diseases [
10]. In contrast, weight gain in men usually accumulates more VAT, resulting in higher metabolic risk [
9].
In subjects with overweight and obesity, changes in WAT depots and cellularity, with a prevalent pro-inflammatory adipokine secretion profile, may directly contribute to the development of metabolic and cardiovascular consequences of obesity [
6,
9].
Adiponectin and leptin are the two most widely studied adipocyte-derived factors. Most leptin is secreted by SAT [
11,
12], flows into the bloodstream, passes through the blood brain barrier and arrives in areas of the brain involved in regulating hypothalamic energy balance. The link between neuroendocrine and sympathetic control of WAT endocrine function, and the existence of negative feedback between the brain and WAT, have been widely studied [
13]. In the last few decades, several studies have shown the association between leptin and cardiovascular diseases [
14].
Adiponectin is an adipokine with described anti-atherogenic, anti-inflammatory and insulin-sensitizing properties [
15]. Obesity is associated with reduced adiponectin expression in VAT, and adiponectin levels have been shown to negatively correlate with the amount of VAT [
16], suggesting that the obesity-associated decline in adiponectin could contribute to the detrimental effects of excessive VAT accumulation on whole body metabolism [
17].
VAT expansion also triggers other proinflammatory cytokine expressions as well as the recruitment of immune cells [
18]. Expansion of VAT is accompanied by increased interleukin 6 (IL-6) and tumor necrosis factor (TNF) secretion, which lead to a crown-like structure formation, and augmented hypoxia-inducible factor 1α expression to promote angiogenesis, which contributes to local and systemic inflammation [
19,
20,
21].
Moreover, great attention has recently been focused on WAT contribution in inter-organ communication, not only by producing signaling mediators but also by converting or degrading signaling mediators from other organs (“signal metabolism” and “signal catabolism”) [
6]. Increasing and specific interest has been given to Evs (carrying protein, lipids, small coding and noncoding RNAs), which are now considered an eminent way of communication between WAT and other organs, as well as between different cell populations within WAT itself [
6].
3. Adipose Tissue Changes across Aging
With aging, quantity, distribution, and function of WAT changes. Fat mass increases and reaches its peak at about 65–75 years for men, and later for women [
22]. The increase in fat mass is independent from changes in body weight, and this is due to the concurrent decline in muscle mass: so-called sarcopenia [
23]. Fat storage is progressively redistributed from the body’s periphery (i.e., loss of SAT, in particular, gluteo-femoral SAT) to the abdomen (i.e., increase of VAT), and such abdominal fat accumulation is independent of weight gain [
24,
25]. It has been shown that involuntary SAT loss, in the absence of a negative energy balance, is associated with triglyceride (TG) spillover, which determines ectopic deposition of TG in muscle, liver, bone marrow and heart, contributing to the dysfunction of these organs [
24,
25].
With aging, WAT also becomes dysfunctional, showing an increased profile of pro-inflammatory adipokines produced by adipose cells, greater infiltration of inflammatory cells in WAT, and preadipocytes and adipocyte incompetence, together leading to inflammaging [
26].
In general, serum levels of the majority of adipokines are higher in older than in younger individuals [
24]. The relationship between aging and the endocrine function of WAT is complex to study in humans. As aging is associated with changes in fat mass and its distribution, as well as with high prevalence of metabolic syndrome, insulin resistance and obesity, the effect of aging itself is difficult to isolate. Indeed, age-related VAT increase, obesity and metabolic syndrome are all factors that can induce an increase in inflammatory and a decrease in anti-inflammatory adipokine production.
Older subjects show higher leptin levels [
27], whose activity seems to be reduced, thereby determining a phenomenon called leptin resistance, a phenomenon that is not completely understood in humans [
28].
It has been determined that the amount of serum adiponectin rises as humans age [
29], and higher levels of adiponectin have been found in centenarians [
30]. Although the beneficial metabolic and anti-inflammatory effects of adiponectin have been confirmed by some scientific studies, adiponectin’s role in the elderly is still controversial [
30,
31]. Indeed, a significant positive relationship was found between adiponectin and risk of incident disability and all-cause mortality among the subjects of the Health ABC Study [
32], but this relationship was not significant after adjusting for weight loss and physical performance at baseline.
In an in vitro model of chronological aging of adipocytes, we and other researchers observed that adipocyte secretion of proinflammatory cytokines, such as interleukin-6 and monocyte chemoattractant protein-1, was significantly higher in older than younger adipocytes [
33,
34,
35]. We also found that in vitro aged adipocytes accumulate ROS, increase mRNA expression of key proteins related to the remodeling of the extracellular matrix and increase p53, p21 and p16 expression, compared to younger cells [
34,
35].
Moreover, dysfunction of WAT is also characterized by increased oxidative stress (OS), mitochondrial dysfunction, reduction in vascularization and hypoxia [
36]. The age-related deregulation of WAT can initiate inflammatory cycles with monocyte recruitment and activation of macrophages. The ratio between pro-inflammatory M1 and anti-inflammatory M2 macrophages increases with aging [
37,
38], as well accumulation of CD3+ T and CD8+ T cells and activation of T and B lymphocytes [
39,
40]. Factors secreted by activated macrophages induce the release of fatty acids from adipocytes that inhibit differentiation of pre-adipocytes and cause de-differentiation of mesenchymal progenitors into mesenchymal adipocyte-like default cells [
41]. These processes induce lipotoxicity both in WAT and in other organs, cause cellular stress responses, promote release of inflammatory cytokines, block adipogenesis and determine further release of lipotoxic fatty acid [
42]. WAT in the elderly is also characterized by reduction of adipocyte size and increase of tissue fibrosis [
43].
That adipocyte size is affected by aging was clearly shown by Donato et al., who observed significantly lower adipocyte areas from WAT in older than younger mice [
44] and by the fact that a high fat diet was able to increase adipocyte diameter in young rats but not in old rats [
45]. Furthermore, age-related decline of adipocyte capacity to stock TG has been shown to be related to fat infiltration inside the muscles, as well as to a deposition in muscles of toxic lipids such as ceramides [
45]. Fibrosis, characterized by an increase in connective fiber content, may be due to an up-regulation of collagen protein [
46]. The OS that occurs in WAT during aging causes oxidative damage to lipids, proteins and DNA. Finally, senescent cells are over-represented in aged WAT, providing a source of many pro-inflammatory cytokines and chemokines, impairing the production of extracellular matrix modifying proteases and further promoting the production of ROS [
47,
48].
4. Muscle as Endocrine Organ
Muscle has also been identified as having a secretory/endocrine function [
7]. Cytokines and other peptides produced, expressed and released by muscle cells are called myokines. Proteomics analyses identified over 650 proteins and peptides produced by muscle cells, yet their precise biological role has been characterized only in a minority of cases [
49]. Myokines act in an autocrine, paracrine or endocrine way. In fact, some myokines exert their effect on muscle itself, taking part in muscle hypertrophy and myogenesis, while other myokines are involved in the regulation of energy metabolism. There is now solid evidence that skeletal muscle, through the production of myokines, communicates with other key organs and regulates lipid mobilization from adipose tissue, liver endogenous glucose production, insulin secretion and thermogenesis [
50].
Many myokines are produced in response to the contraction of muscle fibers and may indeed mediate protective effects of physical exercise and counteract the pathological consequences of a sedentary lifestyle.
Several myokines act precisely within skeletal muscle itself and are involved in muscle cell proliferation, differentiation, and regeneration [
51,
52]; others are involved in mediating energy supply during exercise (
Table 1).
It is now believed that every stage of the myogenic process involves regulation by myokines, many of which contribute to myogenic regulation at different stages, from satellite cell proliferation to differentiation and cell survival [
49,
53].
The role of IGF-1 and IGF-2 as endocrine modulators of myogenesis has been extensively studied [
54], as they seem indispensable for the initiation of differentiation [
55]; IGF-1 leads to muscle hypertrophy by activating satellite cells and possibly inhibiting autophagy [
56]. Impairment of IGF-1 signaling has been described in chronic disorders, and as such, represents a possible mechanism in muscle atrophy led by altered protein synthesis, autophagy, and impaired muscle regeneration [
57].
Furthermore, IL-7 plays an essential role in myogenesis and may influence the differentiation of satellite cells into fully developed skeletal muscle cells [
58].
On the other hand, TGF-beta has shown to be a strong inhibitor of myogenic differentiation in vitro [
59]. Myostatin, the first discovered myokine, is a member of the TGF-beta superfamily and plays a key role in muscle growth and differentiation, by controlling the proliferation of myoblasts (as a major negative regulator of skeletal muscle growth) and suppressing satellite cell activation and myoblast proliferation [
60,
61]; it is known that the deletion of the myostatin gene causes massive muscle hypertrophy in animals [
62,
63].
More recently, follistatin and decorin have been identified as potent inducers of muscle hypertrophy with an anti-myostatin function [
64]; in particular, decorin acts in an auto/paracrine manner as a direct antagonist of myostatin [
65]. Although myokine IL-6 is mainly known for its role in the regulation of lipid and glucose metabolism, it has also been shown that it has an anabolic effect in the processes of myogenesis [
66]. Leukemia inhibitory factor (LIF) also exerts an autocrine/paracrine action [
67], and it has proven to be crucial for satellite cell proliferation and survival [
67].
Apart from the regulatory effects on myogenesis, many myokines act on metabolic pathways in the modulation of energy metabolism. IL-6 [
50] and brain-derived neurotrophic factor (BDNF) [
68] are involved in activating fat oxidation in muscle cells. Indeed, IL-6 signaling within muscle cells appears to affect both glucose uptake and fat oxidation, and its role in GLUT4 translocation has been described [
69]. In addition, several studies described an increase in intramyocellular and whole-body fatty acid oxidation in response to myokine IL-6 [
69,
70]. BDNF affects myogenesis through activation of satellite cells [
71], especially in response to muscle injury; it has also been suggested as a regulator of neuromuscular function during the aging process, with possible implications in sarcopenia and SO [
72]. Indeed, low levels of BDNF are described in subjects with obesity and T2D [
73].
Table 1. Selected myokine functions.
| Function |
Myokine |
Aging |
References |
| Myogenesis and muscle hypertrophy |
myostatin
LIF
IL-6
IL-7
IL-15
musclin
follistatin
decorin
myonectin
IGF-1
musclin |
⇑
?
⇑
⇓
⇓
?
⇓
⇓
⇓
⇓
? |
[61,74]
[67]
[66,75]
[58]
[76]
[77]
[64,78]
[79,80]
[81]
[56]
[82] |
| Muscle-cell FFA oxidation |
IL-6
BDNF
irisin
myonectin |
⇑
⇓
⇓
⇓ |
[69,83]
[68]
[84]
[81] |
| Insulin sensitivity |
IL-6
IL-15
SPARC
LIF
BMP-7
mitsugumin 53 |
⇑
⇓
⇓
?
⇓
? |
[7,69]
[85,86]
[87]
[88]
[89]
[90] |
| Osteogenesis |
IGF-1
decorin
IL-6 |
⇓
⇓
⇑ |
[91]
[92]
[93] |
| Browning of WAT |
Irisin
IL-6
meteorin-like
FGF-21
BAIBA
follistatin
myonectin
BMP-7 |
⇓
⇑
?
⇓
⇓
⇓
?
⇓ |
[84]
[94]
[95]
[96]
[97,98]
[99]
[100]
[101] |
| Lipolysis |
IL-6
FGF-21
ANGPTL-4 |
⇑
⇓
? |
[7]
[102]
[103] |
| Muscle innervation |
BDNF
FGFBP-1 |
⇓
? |
[104]
[105] |
| Muscle angiogenesis |
IL-8
VEGF-A |
⇓
? |
[76]
[106,107] |