1. Introduction
In 1957, Crane et al. isolated a new quinone compound, initially referred to as Q-275, from beef heart mitochondria and described its ability to undergo reversible oxidation and reduction [
1]. With its functional quinone moiety and its ubiquitous presence in living cells, this compound was later officially named ‘ubiquinone’ by the IUPAC-IUB Commission on Biochemical Nomenclature [
2,
3]. However, there were other common names for ubiquinone, including Coenzyme Q
10 (CoQ
10), CoQ, ubiquinone-Q10, vitamin Q10, and ubidecarenone [
4].
CoQ
10 consists of two functional groups, i.e., the five-carbon isoprene ‘tail’ and the benzoquinone ‘head’. The lipid-soluble tail consists of ten isoprenes, with a total of 50 carbon atoms, hence the name ‘CoQ
10’ [
5,
6]. Humans can internally synthesize CoQ
10. The synthesis of the benzoquinone head requires amino acids, either tyrosine or phenylalanine, while the mevalonate pathway provides the acetyl coenzyme A for the polyisoprenoid tail synthesis [
5,
7,
8]. The CoQ
10 head and tail are synthesized in the cytosol but the joining of both functional groups occurs in the mitochondria [
5,
9]. The CoQ
10 biosynthesis requires support from various macro- and micronutrients, such as pantothenic acid for the CoA precursor, pyridoxine for the benzoquinone production, and s-adenosyl methionine for methylation support and isoprene production [
10].
Humans can internally synthesize CoQ
10, while the dietary CoQ
10 sources additionally contribute to its total tissue pool. The main dietary sources of CoQ
10 include fish and meat, while vegetables generally contain low CoQ
10 quantities. Dietary CoQ
10 sparsely absorbs in the hydrophilic intestinal environment due to its lipophilic and high molecular weight properties. Following the gut uptake, CoQ
10 circulates in the lymphatic system and ultimately drains into the blood circulation [
11]. Consequently, the CoQ
10 level in the liver and plasma lipoproteins appears shortly after absorption, but the elevation of plasma CoQ
10 level varies by the blood cholesterol and lipoprotein concentrations [
6,
12]. The CoQ
10 turnover in the body is relatively fast, with a 49 to 125 h half-life, depending on the tissue type [
13].
The combination of endogenous biosynthesis and dietary intake is largely sufficient to prevent the CoQ
10 deficiency state in a healthy individual [
14,
15]. However, cumulative studies reported the association of low CoQ
10 levels in specific conditions such as following strenuous exercise, during aging, after taking some prescribed medications, in patients with various metabolic disorders, and in individuals with cancers [
6,
8,
16,
17,
18,
19,
20,
21,
22]. Despite the documented low CoQ
10 levels, the clinical outcomes of CoQ
10 interventions, either single or in combination with other nutrients, were generally inconsistent.
2. Physiological Roles of CoQ10 in Humans
2.1. CoQ10 Roles in Mitochondrial Bioenergetics
CoQ
10 roles are crucial to the mitochondrial respiratory chain as the electron acceptor. It modulates the electron transferring from the bioenergetic-derived reducing equivalents, i.e., nicotinamide adenine dinucleotide (NADH) and flavin adenine dinucleotide (FADH
2), through the complex I, II, and III in the electron transport chain (ETC). The concurrent proton flow, from the mitochondrial matrix to the intermembrane space, generates the intermembrane proton gradients, which are essential for oxidative phosphorylation and subsequent adenosine triphosphate (ATP) synthesis [
5]. Accordingly, CoQ
10 intervention could have potential bioenergetic benefits in clinical conditions with mitochondrial dysfunction.
While mitochondria are the bioenergetic hub of the cells, they are also the predominant source of reactive oxygen species (ROS) production, oxidative stress, and immunologic and apoptotic regulation. The imbalances of these physiological processes underly diverse metabolic conditions [
25]. As the critical supporter of mitochondrial functions, the importance of CoQ
10 might extend beyond bioenergetics.
2.2. CoQ10 Role as an Antioxidant
With its reversible redox potential and membrane-associated locations, CoQ
10 renders antioxidant protection to the organelles and cell lipid membranes [
13,
26,
27,
28]. Inside the cells, the benzoquinone head of CoQ
10 exists in three interchangeable oxidation states, i.e., the fully reduced ubiquinol (CoQ
10H
2), the ubisemiquinone intermediate (CoQ
10H•), and the fully oxidized ubiquinone (CoQ
10). These redox states are culpable for the scavenging of ROS as well as the mediation of electrons transferring in the mitochondrial ETC.
Nevertheless, the integrated redox modulation of CoQ
10 requires support from other nutrients, specifically α-tocopherol, vitamin C, and other micronutrients.
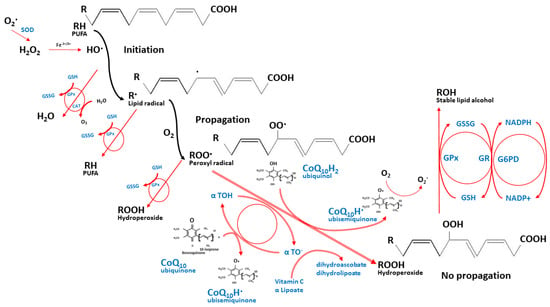
Figure 2 depicts this integrated antioxidant network of CoQ
10 against lipid peroxidation [
29,
30]. While the ROS induces unsaturated lipid peroxidation, it yields the highly reactive lipid peroxyl radicals, which are quickly neutralized by α-tocopherol through the donation of its hydrogen to the peroxyl radicals, thus holding their propagations within membranes and circulating lipoproteins. The reduced ubiquinol then helps regenerate the α-tocopherol antioxidant capacity through their redox interactions. Thereafter, the ubisemiquinone intermediate can either react with the oxygen molecule and produce the superoxide anion radicals or oxidize further to the fully oxidized ubiquinone that does not react with oxygen. Reduced NADP (NADPH), glutathione, and other antioxidants such as vitamin C then help to regenerate the oxidized ubiquinone and α-tocopherol back and maintain their reduction states [
13,
31,
32,
33]. In this integrated manner, CoQ
10 limits the production of lipid peroxyl radicals and protects the circulating lipoproteins, the cellular membrane proteins, the mitochondrial DNA, and the ETC membranes [
26,
34,
35,
36].
Figure 2. The integrated antioxidant defenses against lipid peroxidation. The nutrient network halts the propagation of lipid peroxyl radicals while their redox interactions maintain their reduction states. These nutrients include coenzyme Q10, α tocopherol, vitamin C, α lipoic acid, glutathione, and the micronutrients that support the activities of antioxidant enzymes such as selenium, manganese, copper, and zinc (not shown in the figure). O2•—superoxide anion radical, H2O2—hydrogen peroxide, Fe2+/3+—ferrous or ferric iron, PUFA—polyunsaturated fatty acids, GSH—reduced glutathione, GSSG—glutathione disulfide, GPx—glutathione peroxidase, CAT—catalase, H2O—water, O2—oxygen, α TOH—reduced α tocopherol, α TO•—oxidized α tocopherol.
For oral CoQ
10 supplementation, a 2020 meta-analysis of 17 randomized clinical trials (RCT) documented its antioxidant potentials, comprising the reduction of membrane oxidative damage level, the enhancement of total antioxidant capacity, and the activation of antioxidant defense system enzymes [
30]. As a dietary supplement, the antioxidant capability of CoQ
10 might provide benefits to clinical conditions with underlying oxidative stress pathophysiology.
2.3. Other Physiological Roles of CoQ10
CoQ
10 also serves as the structural component of the ETC membrane supercomplexes that ascertains the efficient ETC functions and prevents the leakage of the electron from the respiratory chain [
28,
37,
38,
39]. The combined result of its structural contribution, lipid peroxidation protection, ROS scavenging, and uncoupling protein activations contribute to the pivotal role of CoQ
10 in mitochondrial membrane integrity [
13,
40]. Besides this, the conservation of mitochondrial membrane permeability is also crucial for cellular survival and functions [
13,
41,
42].
Apart from the mitochondria, the containment of highly acidic enzymes within the lipid-membrane boundary of lysosomes requires the CoQ
10-induced intermembrane proton gradient [
8,
43]. Furthermore, CoQ
10–redox interaction maintains the balance of cytosolic redox intermediates such as NADH, NADPH, and FADH
2. The CoQ
10-mediated reaction also supplies orotate for the de novo pyrimidine synthesis through the oxidative activity of dihydroorotate dehydrogenase [
44]. These intracellular redox balances influence several cellular signalings and gene transcriptions. Such homeostasis modulates apoptosis, bioenergetics, cell growth, and inflammatory responses [
28,
45].
Interestingly, the oral CoQ
10 supplementation showed different 115 gene expressions in the muscle tissue samples from aged individuals compared to their placebo controls [
46]. These findings supported the diverse physiologic roles of CoQ
10 and the potential benefit of its intervention. Several clinical studies also reported that oral CoQ
10 supplementation showed anti-inflammatory effects, including the reduction of tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and C-reactive protein (CRP) [
47,
48,
49,
50]. This immunomodulatory potential of CoQ
10 supplementation also suggests its potential benefit to various immune-mediated clinical conditions.
3. Potential Roles of CoQ10 supplementation in specific medical conditions
Despite the ubiquitous endogenous biosynthesis, specific medical conditions are associated with low circulating CoQ10 levels. However, the inconsistent results of CoQ10 interventions, from previous studies, implied the presence of unaccounted factors that contributed to clinical outcomes [1]. After reviewing the participants’ status in previous CoQ10 clinical trials, we herein proposed two potentially confounding aspects, i.e., differences in host metabolic status and the need for CoQ10 interacting nutrients.
Human metabolism fundamentally relies on host macro- and micronutrient reserves. Depleted host nutrient reserve leads to metabolic triage of nutrients toward the preservation
of short-term metabolic survival, usually at the cost of compromised long-term health [224,225]. The protein deformations, with altered enzyme binding constants for various coenzymes, underly these nutrient triage processes [226]. Compromised host nutrients induce metabolic triage and accelerate the pathophysiologies of degenerative
and metabolic diseases [227]. Therefore, nutrient interventions could hinder mitochondrial decay and delay age-associated illnesses [225].
Even the so-called healthy subjects were still prone to conditional micronutrient inadequacies following intense physical activities, despite their good metabolic statuses and no established clinical diagnosis at baseline. The combined nutrient interventions hold better chances to address the conditional nutritional insufficiencies than a single nutrient.
To support this notion, previous studies showed the favorable trends of combined CoQ10 intervention in healthy adults and athletes performing exercise sessions, as compared to
single supplementation.
Elders and patients with diagnosed metabolic and immune-related disorders likely had compromised metabolic status, along with multiple nutrient insufficiency. The depleted
nutrient reserves increased with the advancement of these chronic situations. Hosts with specific nutrient depletion, such as primary CoQ10 deficiencies, benefited from single
CoQ10 supplementation, even though the outcomes were mostly palliative, not curative. The single intervention was also beneficial in hosts with early stages of declined metabolic
status, such as the elderly. The benefits decreased with the advancement of metabolic conditions, as seen in patients with diabetes, cardiovascular diseases, or kidney failure.
Despite the improvement of some surrogate biomarkers such as proinflammatory cytokines, antioxidative capacities, and lipid or glycemic profiles, the positive trends in clinical
outcomes were less promising with single supplementation. Contrarily, combined CoQ10 interventions provided more encouraging results in hosts with impaired metabolic status
due to the readily available interacting nutrients in the formulations.
However, both CoQ10 interventions would be less beneficial in far-advanced conditions with established damage such as neurodegenerative conditions or cancers. The results
of both single and combined supplementation, at best, affected some surrogate biomarkers but not the overall clinical outcomes. Therefore, nutrient interventions are preventive or
protective rather than curative measures.
In conclusion, no single nutrient could magically drive whole physiological processes. Single CoQ10 supplementation will be beneficial only for hosts that specifically require it, such as hereditary CoQ10 deficiencies. The single intervention will be less promising when the host metabolic status worsens with the likelihood of multiple nutrient insufficiencies. On the contrary, the mixed CoQ10 supplementation with other interacting nutrients will create more promising impacts in hosts with compromised nutrient reserves. However, the results of either single or combined intervention will be less promising in far-advanced conditions with established damage.
With the limited amount of high-level evidence, such as provided by systematic reviews and meta-analyses, we could only conclude that the considerations of whether
to take supplementation varied by the individuals’ metabolic status and their nutrient reserves, which span across the continuum of metabolic triage processes that lead to chronic
health issues. Future studies are warranted, particularly for the RCT with the design to control the host metabolic and nutrient status of participants and the meta-analysis of
upcoming CoQ10 studies on each subject’s metabolic status.
This entry is adapted from the peer-reviewed paper 10.3390/nu14204383