Cultivated tomato,
Solanum lycopersicum [
1] (old name:
Lycopersicon esculentum Mill), originated and domesticated in Western South America and Central America from Mexico to Peru [
2]. Tomato is a member of division Magnoliophyte, class Magnoliopsida, sub-class Asteridae, order Solanales, and family Solanaceae. Tomato is unquestionably the most popular garden crop worldwide [
3], botanically classified as fruit (berry) but often consumed as vegetable (raw, cooked and/or in processed form), next to potato [
4,
5]. Although the cultivated tomato is a tropical plant, it requires moderate growing conditions and is widely adapted to different climates, thus making it cosmopolitan. China, USA, India, Turkey, Egypt, and Italy are some of the major tomato-producing countries [
5]. India ranks third among the top tomato-producing countries, and it is cultivated in 6.34 million ha, with a total production of 12,433,200 tonnes and average productivity of 19,598 kg/ha [
5]. The largest tomato-producing states in India include Maharashtra, Bihar, Karnataka, Uttar Pradesh, Odisha, Andhra Pradesh, Madhya Pradesh, and Assam [
6]. Worldwide, more than a thousand cultivars of tomato are grown, selected based on fruit size, shapes, and growth pattern in different environments [
3]. Plants of tomato are diploid (2n = 2x = 24), determinate to indeterminate growth habit, herbaceous with bisexual flowers, annual to perennial, self-pollinated, and are commonly propagated by seeds. Generally, seedlings of 4–5 weeks old are transplanted, prior to which hardening should be performed by withholding water for up to 4–5 days [
7,
8]. With proper water supply, tomato can be cultivated in different soil types, such as clay, black soil, and red soil; nonetheless, organic-matter-rich sandy loam soil is ideal. Tomato can also tolerate moderate saline and acidic soils. Temperature of 15–27 °C is found to be optimum for its cultivation; however, day temperatures exceeding 38 °C may adversely affect fruit set. In general, tomato is a self-pollinated crop; however, 30% cross-pollination has been recorded in some cases [
8,
9]. Most tomato genotypes have compound leaves, pinnately dissected with 5–9 leaflets on petioles. The inflorescence is a cyme with monochasial or dichotomous branching patterns. Flowers are bisexual and usually yellow in colour; the male part of flower is the stamens, consisting of anther borne on a filament. Anthers join laterally to form a flask-shaped cone with a prolonged sterile tip at the apex. The female flower part is the pistil, consisting of stigma, style, and ovary, and is situated in the flower’s centre, encircled by the stamens. Though tomato is commonly classified as a vegetable, it is really a fruit; berry. Most cultivated varieties, except cherry tomatoes, are either bilocular or multilocular with 4–5 locules [
10]. Fruits come in a variety of shapes (globose, round, oval, flattened, elongated, or heart-shaped) and colours (golden, yellow, red, pink, purple, green, striped to white) [
11]. Tomato fruits are rich in vitamin A, vitamin C, vitamin E, potassium, dietary fibre, β-carotene, and lycopene. Upon ripening, fruits develop a characteristic deep-red colour due to the pigment lycopene [
12]. Among dietary carotenoids, lycopene is one of the most powerful antioxidants that protects against free radicals and is also known to prevent cancer [
12]. Besides its nutritional and therapeutic properties, tomato is used for its distinct flavour and as a food colourant. Tomato fruits are also used to make a range of value-added products, such as whole-peeled tomatoes, diced products, paste, sauce, juice, and soups [
13,
14]. Tomato’s projected genome size is 900 megabases (Mb), containing 34,727 protein-coding genes [
15]. Genes are mostly located in the euchromatin region, which accounts for less than a quarter of total genomic DNA [
16]. Relatively small genome size makes tomato a genetic model for crop improvement [
8] and, also, a suitable system for studying functional genomics, proteomics, and metabolomics. Further, it has been widely used for studies of various biotic and abiotic responses due to availability of various resistant genes and adaptation to a wide environmental condition and photoperiod insensitivity [
17,
18,
19,
20,
21]. The tomato genome consortium sequenced the whole tomato genome as a key model plant for speeding genomic research in the
Solanaceae family because it has the same haploid set of chromosomes and conserved genome structure (a high level of synteny) as other Solanaceous plants. Tomato was the first transgenic food (Flavr Savr Tomato) crop released for commercial cultivation in 1992 in U.S.A., genetically engineered to increase the shelf-life of fruits [
22]. Being a model crop for molecular study and genetic manipulation, tomato is the highest studied crop among the vegetable crops. To date, numerous transgenes were transferred in tomato and evaluated against the biotic (virus, bacteria, fungi, and insect) and abiotic (drought, salt, heat, cold, and water logging) stress tolerance and quality improvement. Among abiotic stresses, drought is the most important stress. Drought is the inadequacy of water, which restricts tomato growth and yield [
14,
23]. Globally, only 14.51 percent agricultural land is well irrigated and the rest is moderately to highly drought susceptible (FAO, 2012). Drought stress primarily affects the crops by generating reactive oxygen species (ROS), which disturbs the cellular homeostasis [
14,
23]. In order for normal growth and development, crop must constantly adapt to climatic conditions. The water loss management in crop and detoxification of ROS are the key strategies in drought stress management. In tomato, for drought stress management, numerous transgenic tomatoes were developed.
2. Drought Stress
Any altered physiological condition that tends to disrupt plants’ homeostatic equilibrium is termed as stress [
9,
13,
19,
24,
25]. Homeostasis is, in turn, a fixed value for metabolism under standard conditions, which is hardly attained by plants, as most of them are exposed to various types of stresses [
24]. Drought, heat, cold, salt, high temperature and light, oxidative stress, heavy metal toxicity, radiations, and UV light are some of the constant threats to plants worldwide. The effect of biotic and abiotic factors on plants is determined by the quantity, intensity, duration, and methods of application on plants. Furthermore, plants have evolved a wide range of physiological and biochemical changes to fit and adapt to a variety of environmental challenges. Nevertheless, globally abiotic stresses are considered as the primary factor for the crop loss which roughly estimates to ~70% of yield reduction [
2,
24,
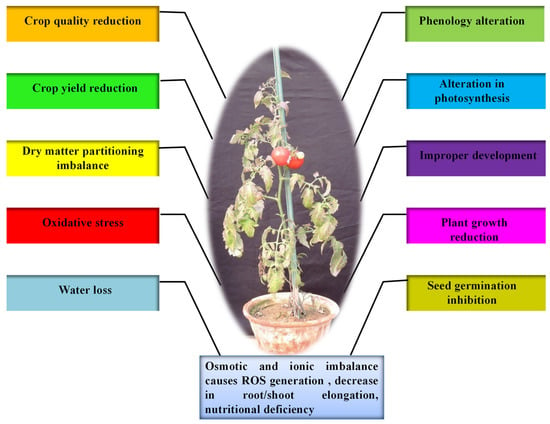
25]. Water deficit is the most critical abiotic stress that affects many physiological parameters of plants, particularly photosynthetic capability. During drought, dehydration of the plant cells and tissues causes diminished plant growth and productivity (
Figure 1). Thus, water deficit works as a limiting factor in agriculture for production by inhibiting the crop from performing its genetic yield potential [
20]. Drought is characterised by inadequate availability of water, due to precipitation variability, evaporation, and soil moisture holding capacity; high water requirement by plants or a combination of all these factors restricts the plants from achieving their full genetic potential. Metrologically, drought is described as a period of below-normal precipitation that limits plant productivity in an agricultural or natural system [
26]. Nearly two thirds of geographical area of India receives low (>1000 mm) or erratic rainfall. Out of 140 million hectares of cultivated area, about 68% is vulnerable to drought stress and ~34% is classified as “severe”, where the drought frequency is nearly periodic. Drought stress affects around 68% of the 140 million hectares of cultivated land, with ~34% categorised as “severe”, indicating the drought frequency being nearly periodic. Among the abiotic stresses, drought is the most severe and pervasive environmental factor [
9,
13,
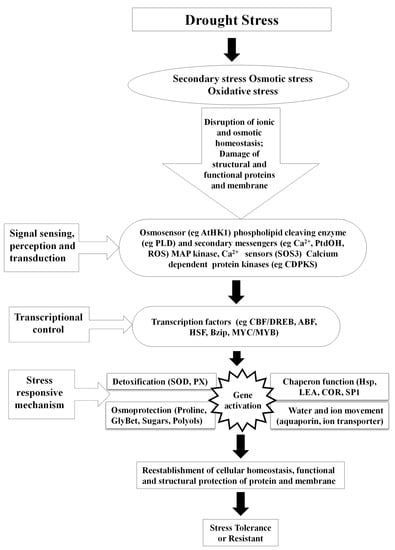
24]. Plants do expedite the detoxification and repair processes in response to stress (
Figure 2), which may include alterations in the expression of ROS scavenging enzymes,
late embryogenesis abundant (
LEA)/
dehydrin-type genes, and the synthesis of molecular chaperones, proteinases, and other detoxification proteins [
9,
27,
28].
Figure 1. The overall impact of drought stress on tomato plant growth.
Figure 2. Mechanism involved in drought stress tolerance.
2.1. Drought-Responsive Transcription Factor Genes in Tomato
Drought stress causes a rise in the levels of endogenous ABA in plants [
29], and a concomitant altered numerous gene expression [
30] in cultivated tomato leaves. Wild relatives of
S.
lycoperscum are resistant to many biotic and abiotic stresses.
S.
pennellii (syn.
L.
pennellii), a wild relative of tomato, grows and reproduces with minimal water, higher water use efficiency, and resistance to wilt [
31]. Martin and Thorstenson [
32] used carbon isotope composition and season-long water use efficiency measures to confirm drought resistance in
S.
pennellii. Kahn et al. [
33] discovered and isolated four tomato genes (
le4,
le16,
le20,
and le25) in response to increased levels of endogenous ABA. Kahn et al. [
33] investigated the expression of these genes in
S.
pennellii to gain a better idea of their function during drought stress. All four ABA-sensitive
S.
lycopersicum genes had homologous counterparts in
S.
Pennellii. In
S.
pennellii, the increase usually occurs after a prolonged duration of drought compared to that in
S.
lycopesocum.
However, only three of the four genes (
le76,
le20, and
le2.5) were expressed in
S.
pennellii in response to exogenous ABA treatment. The expression patterns of these genes in
S.
pennellii were usually parallel to those in
S.
lycopersicum, indicating a similar but undetermined function in both genotypes rather than having genes that are responsible for improved drought tolerance in
S.
pennellii. Members of the abscisic-acid-responsive element binding protein (AREB)/abscisic acid element binding factor (ABF) subfamily of
basic leucine zipper (
bZIP) transcription factors have been involved in the response to ABA and abiotic stress. In
Arabidopsis,
AREB1 and
AREB2 functional evaluation against ABA, drought, and salinity stress conditions are revealed [
34,
35]; however, the tomato
AREB function and transcriptional regulation remain unknown [
36]. Yaez et al. [
37] identified a basic leucine zipper (
bZIP) transcription factor in
Solanum lycopersicum (
SlAREB1),
Solanum chilense (
ScAREB1), and
Solanum peruvianum (
SpAREB1). The three proteins’ deduced amino acid sequences were 97% similar and exhibited strong homology with the
ABF/AREB subfamily of transcription factors which was demonstrated in other plant species, such as
Arabidopsis (
ABF2, 54% similar) and tobacco (
PHI-2, 50% similar). The induced
SlAREB1 expression was reported in
S.
lycopersicum by abscisic acid, cold, and drought. For a quick investigation of genes controlled by
SlAREB1 in tomato and tobacco, a simple transient expression assay using
Agrobacterium-mediated transformation was performed. Tobacco leaves expressing
SlAREB1 exhibited upregulation of stress-responsive genes, such as
rd29B,
LEA ERD10B,
TAS14,
PHI-2, and
trehalose-6-phosphate phosphatase [
37]. These findings imply that
bZIP has a function in abiotic stress response in
Solanum species. In the case of
S.
lycopersicum, [
38] Orellana et al. identified
SlAREB1 and
SlAREB2 are induced in leaf as well as in root tissue by drought and salinity, but the effect on
SlAREB1 was more than that on
SlAREB2. The
SlAREB1 overexpression in transgenic tomato plants improved salt and water stress tolerance compared to non-transgenic tomato plants.
SlAREB1 target gene microarray and
cDNA-amplified fragment length polymorphism (AFLP) analyses revealed that genes coding for oxidative-stress-related proteins, lipid transfer proteins (LTPs), transcription regulators, and LEA proteins were found to be upregulated in
SlAREB1-overexpressing tomato lines, especially in aerial tissue. The findings indicate that
bZIP transcription factor (
SlAREB1) is associated with ABA signal pathway that contributes to abiotic stress, as well as in pathogen response [
38]. Hsieh et al. [
36] found
AREB gene, a
SlAREB transcription factor, using a tomato
cDNA microarray. Electrophoretic mobility shift tests revealed that the chimeric DNA-binding domain of
SlAREB proteins can bind to the promoter regions of
Atrd29A and
SlLAP. However, an ABA-dependent post-translational alteration is required for the SlAREB protein trans-activation [
36].
SlAREB constitutive expression improves tolerance to drought and salt stressors in
Arabidopsis and tomato plants, preserving PSII, membrane integrity, and water availability in plants. Stress-related genes
AtCOR47,
Atrd29A, and
SlCI7-like dehydrin were regulated by
SlAREB overexpression in
A.
thaliana and tomato plants subjected to abiotic and ABA stress. Hsieh et al. [
36] found that
SlAREB regulates some genes involved in stress response and its overexpression improves plant tolerance to drought and salt stress. Islam and Wang [
39] isolated full-length
LeDREB3 cDNA from tomato, investigated the pattern of expression in tomato under various abiotic stresses, and suggested that the
LeDREB3 gene may be responsible for tomato plant stress tolerance.
LeDREB3 is present in duplicate copies in the tomato genome and has considerable sequence match to DREB proteins belonging to various species of plant family. The
LeDREB3 constitutive expression reported in all the tested organs, which was especially higher in flower. Drought, low temperature, salt, and H
2O
2 were shown to probably enhance
LeDREB3 expression, which is expected to provide tolerance to these stresses. However, they did not evaluate its constitutive transgenic expression in tomato or any other plant species.
Investigating the ethylene response factor (ERF) protein demonstrates how it affects the expression of downstream genes in plants under stress by interacting with the GCC box, dehydration-responsive element, and C-repeat [
40]. By inhibiting antisense
TERF2/LeERF2, the ethylene-inducible ERF protein
TERF2/LeERF2 demonstrates the positive control of ethylene production in tomatoes [
41]. According to Zhang and Huang [
40], increased freeze tolerance in tomato and tobacco is associated with the control of
TERF2/LeERF2, and cold gradually triggers the expression of
TERF2/LeERF2. Similar to
Arabidopsis CBF1–
3 genes, tomato encodes
LeCBF1–
3 (
L.
esculentum CBF1–
3), three homologs that are tandemly present within the genome [
42]. However, only tomato
LeCBF1 gene is reported to be induced by cold.
LeCBF1-3 transcripts did increase in response to mechanical agitation, similar to
Arabidopsis CBF1-3, but not in response to ABA, drought, or high salt.
LeCBF1 is a homolog of the functional proteins
CBF1-3 in
Arabidopsis, and its constitutive overexpression in transgenic
Arabidopsis increased the expression of
CBF-targeted genes and improved tolerance to freezing. Tomato harbours an absolute
CBF cold response pathway, according to Zhang et al. [
43]. However, the tomato
CBF regulon differs from that of
Arabidopsis and seems to be substantially smaller and less functionally diverse. A number of dehydration-responsive elements-binding proteins (
DREBs) in plants have been identified, and it has been suggested that both abiotic and biotic stressors can activate them. Recently, Guo and Wang [
44] isolated a
DREB gene from tomato and designated it as
LeDREB2.
LeDREB2 is classified in
DREB family with the
AP2 group. Being a single-copy gene of the tomato genome, it expresses strongly in young leaves and roots but shows weak expression in shoots and mature leaves. Various environmental stresses, such as cold and draught, induce the transcription of
LeDREB2. Different studies demonstrate the expression analysis inducing the transcription of
LeDREB2 gene due to various stresses (H
2O
2, salt, ABA, and methyl viologen) in tomato. Guo and Wang [
44] found that
LeDREB2 gene is a
DREB transcription factor involved with oxidative and abiotic stress in tomato. Recently, Li et al. [
45] cloned
SlDREB from cultivated tomato
M82, a transcription factor, and found that it plays a negative role in architecture of tomato plant, and elevates tolerance against drought. Expression profiles indicate that
SlDREB expression is mainly expressed in leaf and stem and is unable to induce by abscisic acid (ABA) but is suppressed by ethylene and GA. The activity of yeast assay exhibited that
SlDREB exclusively binds to
dehydration responsive element/C-repeat (
DRE/
CRT) of the
SlCPS promoter region. When constitutively overexpressed, the
SlDREB altered plant morphology by regulating leaf and internode elongation, and the consequent dwarfism of tomato plants could be reversed by adding gibberellic acid (GA3) exogenously. The findings of Hsieh et al. [
36] showed constitutive overexpression of
SlAREB in tomato induced dwarfism that could be alleviated by exogenous
GA3 treatment, which were comparable to findings of [
45]. Transcript level analysis of transgenic plants disclosed that
SlDREB overexpression resulted in dwarf phenotype by downregulating key
GA biosynthesis genes involved, such as
ent-copalyldiphosphate synthase (
SlCPS) and GA 20-oxidases (
SlGA20ox1, -
2, and -
4), thereby reducing endogenous GA levels of application in transgenic plants.
2.2. Genetic Engineering of Tomato for Improved Drought Stress Tolerance
In the last century, efforts through classical breeding have resulted in improved agronomic traits, as well as nutritional value of cultivated tomato. However, traditional breeding has not been very successful in enhancing drought tolerance in tomato cultivars due to the limited genetic variation within
S.
lycopersicum species. It is evaluated that just ~5% of the complete genetic variability inside the tomato family can be found within
S.
lycopersicum (100 Tomato Genome Sequencing Consortium, 2014). The genes related to different required agricultural traits, including drought resistance, do not exist in this cultivated species [
13]. However, fortunately, related wild tomato species, including S.
pimpinellifolium,
S.
chilense,
S.
peruvianum,
S.
pennellii, and
S.
hirsutum, have shown to be a rich source of the genes and traits needed to increase resistance to various abiotic stresses. Due to the intricacy of abiotic stress tolerance mechanisms, transferring these traits from wild relatives of
S.
lycopersicum is challenging and takes a lot of effort; therefore, they have not been fully utilised [
13]. The genetic base for resistance and tolerance against abiotic stress in species of wild tomato are hereditary as quantitative traits; thus, it is also unlikely that a single gene from wild tomato species expressed in cultivated species will confer drought tolerance [
46]. For such reasons, developing stress-tolerant transgenic tomato taking regulatory genes from related or distant species is a workable approach to improve drought tolerance of tomato. Therefore, genetic engineering is a relatively quick, precise, and frequently successful method of improving abiotic stress resistance in many plant species.
Considering the problem of drought stress in tomato, attempts have been made to enhance drought tolerance by incorporating single genes from distant plant species [
8,
19,
20,
24,
27,
47], as well as some microbes, governing drought and other water-deficit stress tolerance. Constitutive overexpression of
Arabidopsis CBF1 in tomato brought about upgraded plant resistance to cold, dry season, and salt burdens. In any case, this upgraded resistance included some significant pitfalls, causing decreased plant development and yield [
19]. The transgenic tomatoes showed improved resilience to dry season, cold, and salt stress when a related gene (
CBF1) was expressed using an ABA/stress-inducible
ARBC1 promoter from the barley
HAV22 gene. Additionally, the use of the inducible promoter eliminated the negative effects of the ectopic expression of
CBF1 on plant growth and yield [
19,
27]. By ensuring various proteins, boiling stable proteins (BSP), have been involved in parching resistance against drought stress. Drought stress resistance of transformed tomato plants was attempted with a special 66-kD BSP from
Populus tremula using polyethylene glycol (PEG) study, biomass investigation, proline assay, and electrolyte leakage measurement. These plants displayed slightly increased drought stress tolerance [
48].
S.
lycopersicum was found to contain the basic leucine zipper (
bZIP) transcription factor
SlAREB1 [
37]. Uncomplicated transitory expression analysis was implemented for fast study of genes controlled by
SlAREB1 in tobacco and tomato recommended regarding the group of
bZIP that performs a role in abiotic stress reaction in the
Solanum genus. Zhang et al. [
49] developed marker-free selectable transgenic tomato plants exhibiting increased resistance to drought, cold, and oxidative stresses, which constitutively expressed
AtIpk2b, an inositol polyphosphate 6-/3-kinase gene from
A.
thaliana. Tobacco
osmotin gene driven by
CaMV35S promoter was transferred to tomato and physiological analysis at T
1 and T
2 generations was used to test for resistance against drought and salt stress. Increased resistance to drought and salt stress was observed during NaCl stress and when water was withheld [
47]. Tomato plants were transformed with the
mannitol-1-phosphate dehydrogenase (mtlD; EC1.1.1.17) gene from bacteria, which is controlled by the
CaMV35S promoter. Drought, cold, and salinity resistance study showed that transgenic lines perform a better resistance against abiotic stresses [
50]. The
SlAREB1-overexpressing transgenic tomato plants exhibited increased resistance against drought and salt stress, as analysed by various physiological parameters, such as relative water content, damage by lipoperoxidation, and chlorophyll fluorescence [
38].
By offering the codA gene expressing a choline oxidase from
Arthrobacter globiformis, Goel et al. [
51] modified tomato for increased resistance to salt and water stress with the capacity to assimilate glycine betaine. The
codA transgenic plants demonstrated increased resistance to salt stress during seed germination and ensuing development of young seedlings compared to wild varieties of plants. Developed
codA transgenic plants had uncovered more elevated levels of relative water content, chlorophyll concentration, and proline level. Rai et al. [
19,
24] developed transgenic tomato
cv. Kashi Vishesh (H-86) for improved drought tolerance by using the
BcZAT12 gene coding sequence regulated through
Bclea1a promoter.
BcZAT12 expresses a C
2H
2 type zinc finger protein that is known to confer multiple abiotic stress tolerance to plants. Analysis of RWC, EL, CCI, H
2O
2 and superoxide anion level, and antioxidant enzyme activities suggested that
BcZAT12 tomato transformants had increased levels of drought tolerance. Transgenic plants with the
A.
thaliana transcription factor
ATHB-7 gene inserted displayed reduced stomatal density and increased tolerance to drought stress [
52]. However,
AtATHB-7 was established to be expressed increasingly under drought stress; thus, it performs as a negative growth regulator. The integration of drought-stress-tolerant genes from unrelated species into tomato has been the focus of intense research efforts from scientists all over the world (
Table 1) [
47,
48,
49,
50,
51,
52,
53,
54].
Table 1. Transgenes used for development of drought stress tolerance, their function, and mechanism of action.
| S. No. |
Gene |
Function |
Mechanism of Action |
Reference |
| 1 |
CBF1 |
Stress-inducible transcription factors |
Expression of stress-responsive gene |
[55] |
| 2 |
H1-S, drought induced linker histone |
DNA packaging and organisation of chromosomes in the nucleus |
Modulation of mechanisms related to the stomatal function |
[56] |
| 3 |
AtCBF-1 |
Stress-inducible transcription factors |
Expression of stress-responsive gene |
[57] |
| 4 |
H+-pyrophosphatase |
Facilitate auxin fluxes |
Enhance pyrophosphate-driven
cation transport into root vacuolar fractions |
[58] |
| 5 |
bspA |
Protein protection |
Enhance desiccation tolerance by protecting proteins in membranes and cytosol |
[48] |
| 6 |
coda |
Accumulation of glycine betaine |
Osmolyte accumulation to protect against oxidative damage. |
[59] |
| 7 |
LeNCED1 |
Increase in abscisic acid
(ABA) accumulation |
Stomatal closure and increased water-use efficiency (WUE) |
[59] |
| 8 |
Osmotin |
Stress-responsive multifunctional protein |
Osmotin provides protection via different mechanisms related with programmed cell death |
[47] |
| 9 |
PtADC |
Induce the stress-responsive gene |
Improves dehydration and drought tolerance |
[60] |
| 10 |
DREBs/
CBFs; ABF3 |
Stress-induced transcription factors |
Enhanced expression of downstream stress-related genes confers drought tolerance. |
[19] |
| 11 |
ZAT12 |
Stress-induced transcription factors |
Enhanced expression of downstream stress-related genes confers drought |
[27] |
| 12 |
AtGAMT1 |
Suppress gibberellin |
GAMT1 overexpression inhibited the expansion of leaf epidermal cells. |
[61] |
| 13 |
SlNAC4 |
Stress-responsive transcription factor |
Modulation of ABA-independent signaling networks |
[62] |
| 14 |
GalUR |
GalUR encodes Lgalactono-
1,4-lactone as a precursor of ascorbic acid |
Ascorbic acid detoxifies superoxide anion radical and hydroxyl radical and also plays a crucial role in scavenging
ROS |
[63] |
| 15 |
EgDREB1 |
Enhances the expression of DRE/CRT and DRE/CRT-containing genes |
Elevated level of antioxidative enzymes scavenge the superoxide radical and accumulation of nonenzymatic antioxidants maintains the cell basic structure under drought and cold stress |
[64] |
| 16 |
CcHRD |
AP2/ERF-like tanscritpion factor |
Regulate many pathways involved in stress tolerance |
[65] |
| 17 |
SlMAPK1 |
Encodes for mitogen-activated protein kinases |
SlMAPK1 improves drought stress tolerance by activating antioxidant enzymes, reducing oxidative damage, and modulating transcription of stress-related genes. |
[66] |
| 18 |
SiDHN |
Encodes for Dehydrins (DHNs) commonly
hydrophilin LEA proteins |
Increased chlorophyll a and b, carotenoid and relative water, proline
and soluble sugar content and improve photosynthetic efficiency and suppress the formation of malondialdehyde
H2O2 and O2 |
[67] |
| 19 |
MdSWEET17 |
Sugar transporters |
Enhances accumulation of sugars, such as glucose and fructose, which act as osmoprotestants and carbon source under drought stress |
[68] |
| 20 |
SlSAMS1 |
S-adenosylmethionine synthetase (SAMS) |
SlSAMS1 modulates the production of polyamines and H2O2 and maintains the cellular homeostatasis |
[69] |
| 21 |
AtDREB1A and BcZAT12 |
Encodes for transcription factors |
Independent expression of AtDREB1A and BcZAT12 gene enhances drought tolerance in tomato |
[14,23] |
| 22 |
SlGATA17 |
Improves phenylpropanoid biosynthesis pathway activity |
DNA-binding domain of GATA
TFs regulates many pathways in plants and enhances drought stress tolerance |
[70] |
| 23 |
CsECR |
Encodes for enoyl-CoA reductase (ECR) enzyme, which is involved in biosynthesis of cuticular waxes and catalyses the last step of very long-chain fatty acids (VLCFAs) elongation |
Ectopic overexpression of CsECR increased the contents of total waxes and aliphatic wax leaves and fruits of the transgenic tomato and improves drought tolerance |
[71] |