1. Introduction
The peptide bond is essential to biology. This secondary amide bond is produced by the condensation of the carboxyl group of one amino acid residue with the α-amino group of the following residue and results in the polymerization of amino acids to produce polypeptides. This central reaction is catalyzed by the ribosome and the sequence of the polypeptide chain is defined by the genetic code. In proteins, the peptide bond is sometimes referred to as the eupeptide bond. In addition, peptide bonds can be found in non-ribosomal peptides, which are secondary metabolites such as antibiotics produced by non-ribosomal peptide synthases [
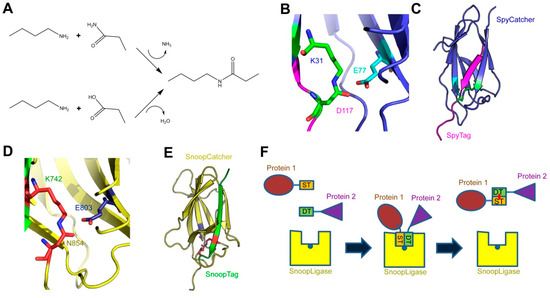
1]. A minority of proteins contain a second class of peptide bonds, referred to as isopeptide bonds. This is an amide bond similar to the peptide bond, but it is produced by the condensation of an amine group with a carboxyl group or a primary amide group located in amino acid side chains, rather than the main chain-building amino or carboxyl groups (A).
Figure 1. The SpyCatcher-SpyTag, SnoopCatcher-SnoopTag, and SnoopLigase systems. (
A) Formation of an isopeptide bond. The primary amine of a lysine side chain condenses with either the side-chain amide of an asparagine (upper reaction) or the side-chain carboxyl of an aspartate (lower reaction) to produce the isopeptide bond, releasing either ammonia or water, respectively; (
B) the isopeptide bond between SpyCatcher-SpyTag. The isopeptide formed by the reactive lysine (K31) in SpyCatcher and aspartate (D117) in SpyTag is shown in green, and the catalytic glutamate (E77) in cyan; (
C) crystal structure of SpyCatcher-SpyTag. SpyCatcher is in blue and SpyTag is in magenta. The structures shown in panels b and c are based on the Protein Data Bank (PDB) entry 4MLI [
20]; (
D) the isopeptide bond between SnoopCatcher-SnoopTag, formed by lysine (K742) in SnoopTag and the asparagine (N854) in SnoopCatcher, is shown in red, and the catalytic glutamate (E803) is in blue; (
E) crystal structure of SnoopCatcher-SnoopTag. SnoopCatcher is in yellow and SnoopTag in green. The structures shown in panels d and e are based on the RrgA D4 domain structure (PDB ID: 2WW8) [
15]; (
F) schematic of the function of SnoopLigase. A protein (in brown) containing SnoopTagJr (ST, in orange) and another protein (purple) with DogTag (DT, green) are mixed in the presence of SnoopLigase (yellow). The tags bind to SnoopLigase, which contains the catalytic glutamate (blue dot) and catalyzes the formation of an isopeptide between SnoopTagJr and DogTag (red dot). The ligated proteins can then be eluted from SnoopLigase.
In eukaryotes, the most prevalent example of isopeptide bond formation is the ubiquitination of proteins targeted for degradation by the 26S proteasome [
2]. The isopeptide is formed by the C-terminal carboxyl group of ubiquitin and the side-chain amine of a lysine residue in the target protein. This reaction is catalyzed by E3 enzymes, or ubiquitin-protein ligases, which also impart specificity to the system [
3]. Similarly, members of the small ubiquitin-like modifiers (SUMO) family, which regulate a number of cellular processes, are ligated to their target proteins via an isopeptide bond [
4].
In bacteria, isopeptides are mostly known from surface proteins of Gram-positive bacteria. Two classes of proteins have notable isopeptide bonds: the microbial surface components recognizing adhesive matrix molecules (MSCRAMMs) and pili [
5]. In pili, pilin subunits are covalently bound to each other via an intermolecular isopeptide bond between the carboxyl group of a C-terminal threonine and the ε-amine of a lysine in the next subunit, a reaction catalyzed by a pilin-specific sortase [
6]. In MSCRAMMs and some pilins, isopeptides are formed spontaneously between residues within the same domain. These domains belong to the immunoglobulin fold superfamily. Isopeptide bond formation increases the stability of the proteins or pili and may contribute to stronger binding to ligands by MSCRAMMs, since in some cases the isopeptide only forms upon ligand binding to lock the protein in a bound conformation [
7].
The SpyCatcher-SpyTag system was developed by the Howarth laboratory based on the internal isopeptide bond of the CnaB2 domain of FbaB, a fibronectin-binding MSCRAMM and virulence factor of
Streptococcus pyogenes [
8,
9]. An internal isopeptide bond forms spontaneously in this domain between the ε-amine of lysine K31 and the side chain carboxyl of aspartic acid D117. The reaction is catalyzed by the spatially adjacent glutamate E77 (B). The resulting isopeptide bond confers high stability to the CnaB2 domain [
10]. The CnaB2 domain can be stably split into two components: a larger, incomplete immunoglobulin-like domain (termed SpyCatcher) of 138 residues (15 kDa) and a shorter peptide (SpyTag) of 13 residues (see for peptide sequences). SpyCatcher contains the reactive lysine and catalytic glutamate, whereas SpyTag includes the reactive aspartate. The two components can still recognize each other with high affinity (0.2 µM) and the isopeptide can form between SpyCatcher and SpyTag to form a covalently bound complex (C). Under experimental conditions relevant to life science research (room temperature, dilute protein concentrations), the reaction rates allow the bonds to form at high efficiency within minutes [
9]. SpyTag in particular is equivalent in size to a number of epitope tags and can be genetically fused to a number of proteins and is able to react with SpyCatcher when inserted at the N- or C-termini of target proteins, as well as internal sites [
9,
11]. SpyCatcher itself can also be produced as a fusion protein, allowing the formation of covalently bonded protein partners that might otherwise be difficult to produce as protein fusions [
12,
13,
14].
Table 1. Summary of Catcher-Tag technologies and their development.
|
Catcher
|
Tag
|
Tag Sequence
|
Description
|
Publication Year
|
Reference
|
|
SpyCatcher
|
SpyTag
|
AHIVMVDAYKPTK 1
|
Original Catcher-Tag technology.
|
2012
|
[9]
|
|
SpyCatcher
ΔN1ΔC1
|
SpyTag
|
AHIVMVDAYKPTK 1
|
Minimal SpyCatcher construct that still binds efficiently to SpyTag.
|
2014
|
[20]
|
|
SpyLigase 2
|
SpyTag
|
AHIVMVDAYKPTK 1
|
Rationally engineered system for ligating two peptides.
|
2014
|
[18]
|
|
KTag
|
ATHIKFSKRD
|
|
SnoopCatcher
|
SnoopTag
|
KLGDIEFIKVNK 1
|
Orthogonal technology to SpyCatcher.
|
2016
|
[16]
|
|
SpyCatcher002
|
SpyTag002
|
VPTIVMVDAYKRYK 1
|
Improved SpyCatcher-SpyTag system with faster reaction rate.
|
2017
|
[21]
|
|
SnoopLigase 2
|
SnoopTagJr
|
KLGSIEFIKVNK 1
|
Rationally engineered system for ligating two peptides.
|
2018
|
[19]
|
|
DogTag
|
DIPATYEFTDGKHYITNEPIPPK
|
|
SpyDock
|
SpyTag002
|
VPTIVMVDAYKRYK 1
|
Protein affinity purification system (Spy&Go) based on SpyCatcher.
|
2019
|
[22]
|
1 The reactive residues are shown in bold. 2 SpyLigase and SnoopLigase have two target peptides.
A similar system has been developed based on another Gram-positive surface protein, the pilus adhesin RrgA of
S. pneumoniae [
15,
16]. The D4 domain of this protein is stabilized by an isopeptide forming between a lysine (K742) and an asparagine (N854), catalyzed by the spatially adjacent E803 [
15] (D). This domain was split into a scaffold protein called SnoopCatcher and a 12-residue peptide termed SnoopTag, which can spontaneously form a covalent isopeptide bond upon mixing [
16] (E). In contrast to SpyCatcher-SpyTag, the reactive lysine is present in SnoopTag and the asparagine in SnoopCatcher. This system is orthogonal to SpyCatcher-SpyTag; that is, SnoopCatcher does not react with SpyTag and SpyCatcher does not react with SnoopTag. This allows the use of both systems simultaneously to produce “polyproteams,” programmed modular polyproteins, for use in biotechnological applications [
16].
The Howarth lab further modified these technologies by making tripartite systems, where the isopeptide-forming lysine and aspartate/asparagine are located on separate peptides, and the catalytic glutamate is present on a larger scaffold protein [
17]. This was first attempted with the SpyCatcher system to produce SpyLigase. Here, a second peptide containing the reactive lysine (KTag) was separated from SpyCatcher, which itself was modified to produce the stable SpyLigase protein containing the catalytic glutamate [
18]. When SpyLigase was mixed with two proteins containing one of the reactive peptides each, SpyLigase was able to catalyze the fusion of the two tags. However, although SpyLigase could mediate the fusion of KTag and SpyTag located at both N- and C-terminal and even internal positions, the reactions had a ~50% efficiency at best and were dependent on specific buffer conditions and low temperature [
16]. In contrast, the recently developed SnoopLigase system appears more robust and efficient [
19]. SnoopLigase was engineered similarly to SpyLigase, and catalyzes the isopeptide formation between the lysine of a modified SnoopTag (SnoopTagJr) and the asparagine in a second peptide termed DogTag (F). This system can have efficiencies over 95% that are less sensitive to temperature and reaction conditions than SpyLigase. Furthermore, as SnoopTagJr and DogTag have relatively high affinity for SnoopLigase, immobilizing SnoopLigase allows washing away unconjugated reactants followed by elution of essentially pure fusion products [
19]. The various Catcher-Tag systems and their development are summarized in .
2. Applications of the SpyCatcher-SpyTag System
Being a quick and reliable coupling tool for irreversible peptide-protein ligation, the SpyTag-SpyCatcher system is ideal for a wide range of applications, ranging from increasing protein stability to antigen delivery during vaccination [
14,
23]. Before focusing on the use of the SpyTag-SpyCatcher system in the investigation of bacterial virulence factors, we give a short overview of those applications. For more detailed information, a list of publications and patents using the SpyTag-SpyCatcher and related technologies is accessible at the SpyInfo web page (available online:
https://www.bioch.ox.ac.uk/howarth/info.htm) and the corresponding sequences and expression routes are listed in the SpyBank database [
24].
A major application of the SpyTag-SpyCatcher system is the formation of so-called SpyRings in order to increase the intrinsic resilience of proteins to denaturation. SpyRings are generated by circularization of a single protein, which is accomplished by fusing an N-terminal SpyTag with a C-terminal SpyCatcher, or vice versa (A). Enzyme circularization increases resistance to hyperthermal denaturation and aggregation, as well as alkali tolerance of individual enzymes without a loss in enzymatic activity [
25,
26,
27,
28,
29]. Enzymes with a short distance between termini (<15 Å), and an active site which is not in direct proximity to one of the termini, are considered ideal candidates for protein circularization [
30]. However, efficient SpyRing circularization and increased thermal stability have been shown for proteins where the termini are even farther apart [
28].
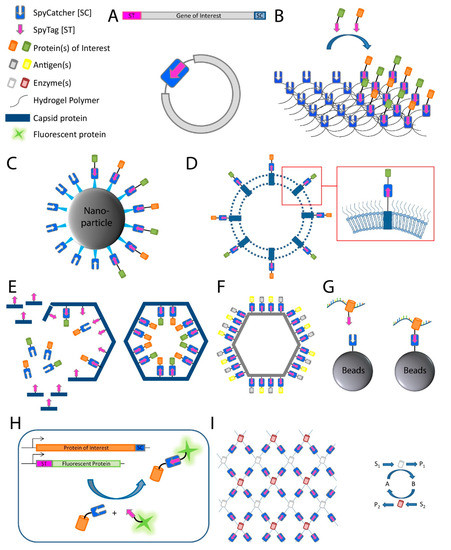
Figure 2. Applications of the SpyCatcher-SpyTag system. (A) SpyRing: SpyCatcher and SpyTag are fused to the terminal ends of the protein of interest resulting in protein cyclization and thereby conferring an increased resilience to denaturation; (B) post-hydrogelation decoration of protein hydrogels: SpyCatcher is fused to the polymeric material used in hydrogel synthesis, which allows the posthydrogelation decoration with proteins of interest fused to SpyTag; (C–F): (C) bioconjugation of target proteins to nanoparticles, (D) outer membrane vesicles, (E) phage capsid proteins to create a proteinaceous nanocompartment, and (F) virus-like particles; (G) SpyCLIP: SpyTagged RNA-binding protein interacts with RNA and is covalently attached to beads with fused SpyCatcher for use in pull-down assays; (H) fluorescent protein labeling for use, for example, in microscopy (ST represents SpyTag sequence); (I) artificial multi-enzyme nanodevices for increased efficiency, stability, and reusability. Schematic of mesh-like nanodevice is shown on the left and an enzymatic reaction scheme on the right. S represents substrate; P represents product; A and B represent cofactors.
Another frequently used application for the SpyCatcher-SpyTag system is the decoration of protein hydrogels (B). Hydrogels have found application, for example as an artificial extracellular matrix material in medicine [
31], tissue engineering [
32,
33], and cell culturing [
34]. Hydrogels are polymeric materials engineered to resemble the extracellular environment of specific tissues with defined functional and structural properties. They have been used for decades as molecule delivery devices and as carriers for cells in tissue engineering due to their ability to mimic aspects of the native cellular environment [
35]. However, hydrogels fail to fully imitate the complexity of biological systems. Fusing SpyCatcher to the polymeric material used in hydrogel synthesis allows the decoration with SpyTagged proteins post-hydrogelation to mimic specific microenvironments [
36,
37,
38,
39]. Thus, the SpyTag-SpyCatcher system is used as a quick and simple molecular tool for the simultaneous incorporation and presentation of different target molecules into and on hydrogels, thereby avoiding an otherwise laborious engineering process.
The SpyTag-SpyCatcher system has also been used for the modular assembly of proteins onto nanoparticles [
40] (C) and bacterial outer membrane vesicles [
41] (D; see also
Section 3.3). The same system can be used for the in vivo encapsulation of enzymes fused to phage capsid proteins in order to create a protein nanocompartment [
42] (E) and for the decoration of virus-like particles (VLPs) for antigen delivery to the immune system [
43,
44] (F; see also
Section 3.3), while successfully preserving the structure and the function of the assembled proteins.
Recently, the SpyTag-SpyCatcher system was used for the characterization of protein–RNA interactions as an alternative for ultraviolet (UV) crosslinking and immunoprecipitation (CLIP) [
45]. The CLIP method relies on the limited specificity of antibody–antigen interactions which cannot withstand harsh washing conditions resulting in insufficient purity of ribonucleoprotein complexes after immunoprecipitation. Therefore, additional gel purification steps are necessary to further purify these complexes, which results in loss of protein. The disadvantages of CLIP are the labor-intensive gel purification steps as well as a high number of false-positive signals due to non-specific interactions. In comparison, the SpyTag-SpyCatcher technology allows the method to be performed with beads, skips the gel purification steps altogether, and withstands harsh washing steps, thus reducing non-specific interactions. The improved method using the Spy technology was termed SpyCLIP and requires SpyCatcher being fused to beads for immunoprecipitation and fusion proteins of the protein of interest, in this case the RNA-binding protein, to SpyTag [
45] (G). UV-crosslinking and affinity purification (uvCLAP) [
46] as well as gel-omitted ligation-dependent CLIP (GoldCLIP) [
47], using the covalent HaloTag-HaloLink purification system for immunoprecipitation [
48], are both improvements to the traditional CLIP protocol, omitting the labor-intensive gel purification steps. The advantage of the SpyCatcher system over those methods is the ability of SpyTag-SpyCatcher to withstand even harsher washing conditions than the biotin-streptavidin coupling used during uvCLAP, and the small size of SpyTag compared to the 33 kDa protein tag used in GoldCLIP.
The system has also found increasing usage in conventional and super-resolution microscopy [
49,
50,
51] by providing an easy and reliable way to label a SpyTagged protein with SpyCatcher coupled to a fluorescent dye (H). Additionally, the limited accessibility of large antibodies, classically used for detection of epitope-tagged molecules, can be avoided by using the relatively small SpyCatcher protein during the detection of SpyTagged targets.
The Spy technology can also be used for the targeting of chemically synthesized voltage-sensitive dyes to specific cells for optical measurement of voltage dynamics in living cells. To this end, SpyTag is linked via a polyethylene linker to voltage-sensitive dyes while expressing SpyCatcher on the target cells. These SpyTag–dye conjugates, termed Voltage Spy, display improved targeting, good voltage sensitivity, and fast-response kinetics [
52].
The system can also be used for the spatial and functional coordination of enzyme functions, creating a microenvironment normally associated with biological compartmentalization and conferring some of the same benefits (e.g., a local increase in enzyme and metabolite concentrations). Linkage of SpyTag with SpyCatcher is utilized for enzyme organization, allowing construction of artificial multi-enzyme nanodevices with a controlled spatial arrangement for increased efficiency of enzyme cascades while maintaining or even increasing stability and reusability of biocatalysts [
53] (I).
3. Using SpyCatcher-SpyTag to Investigate Bacterial Virulence Factors
The SpyCatcher-SpyTag system has recently been used for investigating bacterial virulence factors. So far, these have been limited to studying surface proteins of Gram-negative bacteria, especially autotransporter proteins. Autotransporters, also called type V secretion systems, are a widespread family of secreted proteins from Gram-negative bacteria, and many of these mediate virulence-related functions [
54]. These proteins, which are divided into several subclasses (type Va through type Ve), have an outer membrane-embedded β-barrel domain and an extracellular region or passenger, which harbors the specific activity of each protein. Although SpyCatcher-SpyTag has been used mostly for studying autotransporters, the methods described below would be applicable to other surface-exposed proteins, both in Gram-negative and Gram-positive bacteria.
The cell surface of Gram-negative bacteria is represented by the outer membrane (OM), which is a complex asymmetric lipid bilayer. The extracellular face of the OM is mainly composed of lipopolysaccharides and outer membrane proteins that include several common virulence factors responsible for adhesion, mobility, and secretion, such as autotransporter adhesins, flagella, and type I-VIII secretion systems, among others [
55,
56]. Studying the expression, secretion, migration, and interactions of OM proteins often requires labeling by reporter proteins or other fluorophores. Labeling OM proteins, however, is challenging. Many fluorescent reporter proteins fail to mature in the periplasm [
57]; thus, genetic fusions with fluorescent proteins are limited to a handful of options [
58,
59,
60,
61]. An alternative approach is non-covalent affinity-based labeling by using antibody–reporter protein fusions, affinity tags, or other high-affinity interactions, for example, colicins ColE9 and ColIa for labeling of the vitamin B12 transporter BtuB [
62]. Alternatively, labeling methods using small organic molecules include amine-reactive fluorescence labeling [
63,
64], cysteine-reactive labeling [
65], or site-specific labeling with unnatural amino acids, and tag-specific labeling [
66,
67]. Each of these techniques, however, has shortcomings. Non-covalent labeling requires high (nM) affinities for the label to remain associated with its target for a significant amount of time, which often makes such labeling unsuitable for time-resolved imaging on the minute scale. Small-molecule labeling often requires harsh treatment of the cells with reactive molecules, and site-specific orthogonal labeling requires co-expression of additional OM proteins that can disturb the system. In this section, we present a few recent examples showing how the SpyCatcher system can overcome many of these challenges by covalently labeling OM-bound virulence factors with high specificity, both with the purpose of studying the virulence factors and of exploiting their properties in vaccine development.
This entry is adapted from the peer-reviewed paper 10.3390/ijms20092129