Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Integrative & Complementary Medicine
Modifications in the microbiota caused by environmental and genetic reasons can unbalance the intestinal homeostasis, deregulating the host’s metabolism and immune system, intensifying the risk factors for the development and aggravation of non-alcoholic fat liver disease (NAFLD). The use of probiotics, prebiotics and synbiotics have been considered a potential and promising strategy to regulate the gut microbiota and produce beneficial effects in patients with liver conditions.
- non-alcoholic fatty liver disease (NAFLD)
- non-alcoholic steatohepatitis (NASH)
- prebiotic
- probiotic
- symbiotic
- microbiota
1. Introduction
The accelerated industrialization and urbanization processes have evidenced the exponential increase in ultra-processed food consumption and the predominance of a sedentary lifestyle. This setting has developed an epidemic of obesity, dyslipidemia, type 2 diabetes mellitus (DM2), and Metabolic Syndrome (MS) that are associated with the development of non-alcoholic fatty liver disease (NAFLD) that is the most prevalent chronic liver disease worldwide [1,2,3]. The global occurrence of NAFLD is 25.24%, and the highest prevalence rate of the disease is found in the Middle East (31.79%), followed by South America (30.45%), Asia (27.37%), North America (24.13%), Europe (23.71%), and Africa (13.48%) [1,4].
NAFLD is a term that encompasses a cluster of disorders related to the macrovesicular accumulation of triglycerides within hepatocytes higher than 5%, thus causing steatosis. This process occurs due to a mismatch of regulatory mechanisms involved in lipid metabolism. Some patients develop a more aggressive subtype of the disease characterized by lobular inflammation plus hepatic balloon degeneration, with or without fibrosis, known as non-alcoholic steatohepatitis (NASH) [5,6,7].
Approximately 22.5% of NASH patients will develop hepatocellular carcinoma, and 20% will develop cirrhosis [8,9,10,11]. This progressive feature makes NASH the second most prevalent cause of liver transplantation in the USA [12]. Besides, NAFLD has an intense correlation to cardiovascular disease (CVD). A meta-analysis conducted by Haddad et al. showed that the prevalence of cardiovascular events in patients with NAFLD (14.9%) was more than two-fold compared to patients without NAFLD (6.2%) [13].
The burden of NAFLD is increasing worldwide, and its complications are severe. Numerous therapies have emerged to treat or slow the disease’s advance. Some studies have recently shown favourable results with interventions carried out in the intestine that reflect improvements in the liver, such as the bio modulation of intestinal microbiota [14,15,16,17,18]. The use of probiotics, prebiotics and synbiotics has been considered a potential and promising strategy to regulate the gut microbiota [19,20]. The intestinal microbiota is an intense and dynamic ambient whose composition continually changes. These alterations, caused by environmental and genetic reasons, can unbalance the intestinal homeostasis, deregulating the host’s metabolism and immune system, intensifying the risk factors for the development and aggravation of NAFLD [15,21].
Some reviews investigated the effects of the use of prebiotics and probiotics in NAFLD patients, but most evaluated the effectiveness only on hepatic enzymes [22,23]. Other reviews evaluated the effectiveness of prebiotics or probiotics alone [24,25,26,27]. Only two reviews compared the effects of probiotics, prebiotics and synbiotics on liver enzymes, but not on other risk factors associated with NAFDL and NASH [23,28]. Souza et al. [28] also performed a review comparing the effects of probiotics, prebiotics and synbiotics on NAFLD. Nevertheless, only four trials were included in the review. For these reasons, this systematic review aims to evaluate the effectiveness of probiotics, prebiotics and synbiotics in the management of NAFLD and NASH. To the best of our knowledge, this is the first review to show the effects of probiotics, prebiotics and synbiotics both in NAFLD and NASH.
2. NAFLD and NASH
NAFLD represents a clinicopathological spectrum of liver diseases extending from isolated steatosis fibrosis to cirrhosis and related to hepatocellular carcinoma development. It is defined as the development of steatosis in more than 5% of hepatocytes identified either histologically or radiologically, in the absence of secondary causes such as significant alcohol consumption (<30 g/day for man and <20 g/day for woman), hereditary liver diseases, or viral hepatitis [29]. This disease is comprised of two main entities: NAFLD and NASH. Histologically, the first includes any case characterized by steatosis with minimal or absent lobular inflammation. The second constitutes a more progressive form, characterized by balloon hepatocyte degeneration and diffuse lobular inflammation with or without fibrosis [5]. An increased risk of cirrhosis, hepatocellular carcinoma, and liver-related mortality is associated with NASH, especially when fibrosis is already present. In advanced stages of fibrosis, the mortality rate increases exponentially [30,31].
Although NASH may be suspected in the case of fatty liver and elevated liver enzymes, liver biopsy with histological examination is the only diagnostic method. On the other hand, the diagnosis of NAFLD may be made either by histological examination or by imaging studies that can detect more than 5% of hepatic steatosis. In this sense, ultrasound (US) and computed tomography (CT) are capable of detecting steatosis involving 20% of hepatocytes, and magnetic resonance imaging (MRI) is capable of detecting stenosis in 5% [32,33].
The NAFLD is currently recognized as a hepatic manifestation of MS [4,34] which shares a common pathogenic pathway in insulin resistance. The pathophysiology involves an imbalance between lipid acquisition, mitochondrial fatty acid oxidation, and its export as part of the very-low-density lipoprotein (VLDL) molecule that generates hepatic steatosis [35]. In this way, it is clear that the criteria for MS (dyslipidemia, hyperglycemia, central adiposity, and hypertension) and weight gain will be considered risk factors strongly related to the disease [36].
In NASH, the two-hit proposal has been used for years to explain the pathophysiology of the disease. The first involves insulin resistance that will cause steatosis, and the second is associated with the inflammatory process generated by lipid oxidation. Nevertheless, both hits are insufficient to explain the disease. Thus, a multi-hit theory was proposed, including (besides the phenomena of the old theory) lipotoxicity caused by the accumulation of free fatty acids, cholesterol, and triglycerides, Kupffer cell activation, myeloid cell recruitment, gut microbiota dysfunction, genetic factors, and diet [35,37,38].
Recently, some authors have joined in favor of changing the name from NAFLD to metabolic (dysfunction) associated with fat liver disease (MAFLD). They believe that the term NAFLD has been described as an exclusionary condition. It exists only when other conditions such as viral hepatitis B and C, autoimmune diseases, or alcohol intake above a certain threshold are absent. However, MAFLD is present in about one fourth of the global population, and it coexists with other liver diseases. Another argument resides in the debate about the safe limit of alcohol intake and the challenge about the application of questionnaires that are faithful to the real consumption of this beverage. The third point is that the new term could simplify the stratification of the disease without the dichotomous classification between NASH and non-NASH. Finally, the authors argue that the new term would consider the heterogeneity of fat liver disease which facilitates the selection of phenotypes for clinical trials as well as therapies. The diagnosis of MAFLD is grounded on the identification of hepatic steatosis by histology, imaging, or blood biomarkers, in association with one of the following three conditions: excess adiposity, presence of pre-diabetes or DM2, or evidence of metabolic dysregulation [39,40,41,42]. Figure 1 shows the relationship between the gut microbiome, NAFLD, and NASH.
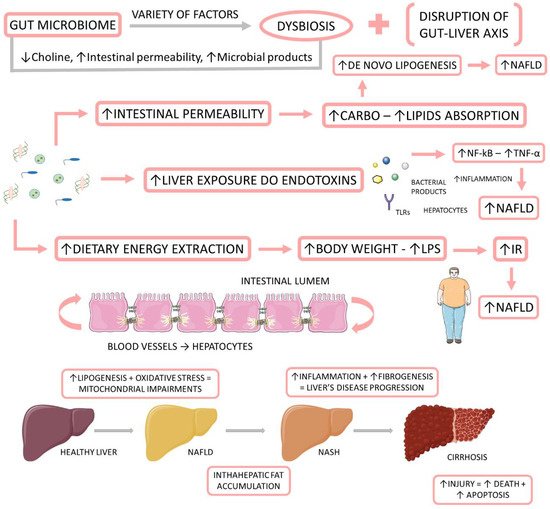
Figure 1. The relationship between the gut microbiome, NAFLD, and NASH. The gut microbiome is affected by a variety of factors to be in dysbiosis. When dysbiotic, the gut microbiome becomes disrupted and starts to cause alterations in the intestinal permeability, leading to augmented liver exposure to endotoxins and dietary energy extraction. These alterations induce an increase in the intrahepatic lipid accumulation, and they induce liver inflammation and fibrosis. IR: insulin resistance; LPS: Lipopolysaccharide; NF-kB: nuclear factor kappa B; NAFLD: non-alcoholic fatty liver disease; NASH: non-alcoholic steatohepatitis; TNF-α: tumor necrosis factor-alpha.
3. Microbiome
The human microbiome refers to the genomic component of organisms (microbiota) that inhabit a specific human body location [43]. More than 30 trillion microorganisms are part of this ecosystem [44]. It is composed mainly of bacteria, but it includes commensal populations of fungi, archaea, and protists as well, covering all three domains (Bacteria, Archaea, and Eukarya), in addition to viruses [45,46]. These microbes reside in the skin, oral cavity, gastrointestinal, respiratory, and genitourinary tracts, accounting for 1–3% of total body weight [47]. Considering the variability between individuals, the intestinal microbiome has a set of 3.3 million different genes, representing a genome 150 times larger than humans’ [48].
The intestinal microbiota plays a vital role in the metabolism of substrates including carbohydrates, proteins, polyphenols, vitamins, and bile [48,49]. It is closely related to the hosts which develop and tune the immune system [50], as well as protect against pathogenic colonization by competing for fixation sites or nutrient sources, producing bacteriocins (e.g., lactic acid), inhibitory metabolites (short-chain fatty acids and lithocholic acid), stimulating IgA epithelization and mucus production [51,52]. Moreover, an interesting relationship between microbiota and the nervous system has been observed, constituting the brain–gut axis (Figure 2). This extensive communication network connects the gastrointestinal tract with the central nervous system’s cognitive and emotional centers (CNS) [53,54].
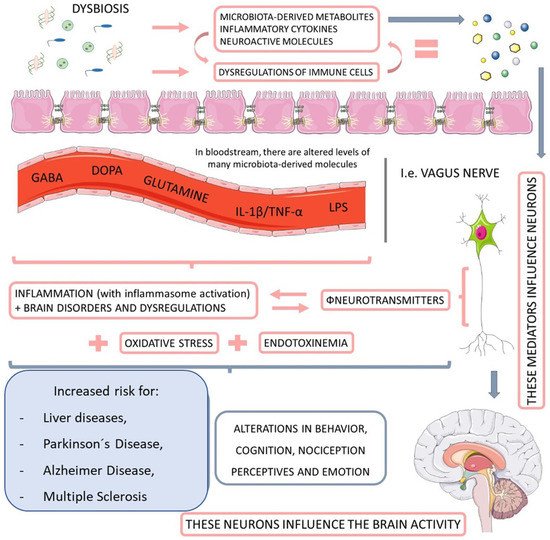
Figure 2. The relationship between microbiota, nervous system, and liver diseases. When dysbiotic, the gut-microbiome-derived metabolites start to cause neuroendocrine dysregulation, principally by impairments in the activity of neurotransmitters. This endotoxemia increases the personal risk for Parkinson’s disease and Alzheimer’s disease, as well as multiple sclerosis and other behavioral and cognitional alterations. GABA: gamma-Aminobutyric Acid; DOPA: dopamine; IL-1ß: interleukin-1β; TNF-α: tumor necrosis factor.
It was established that the fetus’s intrauterine environment was sterile for a long time, and the microbiome development started during and after birth [55]. However, in the last 15 years, with advances in DNA sequencing technology, this theory has been challenged by scientific evidences that demonstrated the presence of microbes in the placenta, amniotic fluid, the umbilical blood cord, and meconium (even in healthy pregnancies), [55,56,57,58,59,60,61] in addition to the possibility of participation of maternal microbiota and its metabolites in fetal development, inciting the theory of in utero colonization [62].
During and after birth, the newborn is surrounded by many microorganisms. Thus, the development and shaping of the initial microbiota will depend on and vary according to factors such as gestational time, mode of delivery, method of infant feeding, intrapartum, and neonatal antibiotic courses [63]. Neonates born from normal births are likely to have fecal microbiota resembling vaginal microbiota, dominated by Prevotella spp. and Lactobacillus, while babies born by cesarean section acquire bacteria derived from the hospital environment and maternal skin, such as Staphylococcus, Corynebacterium, Propionibacterium spp. [64]. This differentiation is relevant as neonates born by cesarean section are more susceptible to developing asthma, rhinitis, food allergy, celiac disease, and overweightness over the years [65]. The mode of infant feeding has a substantial influence because breast milk is rich in human milk oligosaccharides (HMOs) which are involved in pathogenic protection, maturation of the intestinal microbiome, and promotion of intestinal barrier function and maturation of immune cells [66]. Bifidobacterium bacteria are the most related to the positive effects of HMOs and are generally the most abundant among taxons found in infant intestinal microbiota (up to 90%) [67] that have their production two times higher in breastfed newborns compared to feed formulas [68]. Gestation time is also essential, as premature neonates’ immature intestines may have peristalsis, poor barrier functions, and immunity, which may precede the onset of infection and the inflammatory process by colonization pathogenic bacteria [69]. Although decreasing the colonization of pathogenic bacteria, intrapartum antibiotics administration was also related to the development of necrotizing enterocolitis (NEC). Besides, antibiotic administration in the first six months was correlated with an increased likelihood of developing asthma and obesity [70]. Other factors such as family environment, geographical, and cultural traditions are also documented as influencing infant microbiota [63].
In summary, the first colonizers of the newborn’s intestinal microbiota are usually facultative anaerobes, followed by the accumulation of obligatory anaerobes, including Bifidobacterium, Bacteroides, and Clostridium for the following six months [71]. With weaning and the introduction of solid foods from six months onwards, intestinal microbiota diversity increases, with Actinobacteria and Proteobacteria becoming the dominant components of infant microbiota [72]. At the age of 2.5, the infant microbiota’s composition, diversity, and functional capabilities resemble adult microbiota [73]. This intestinal community will undergo subtle changes until middle age (around 40 years of age), which is a time of relative stability [74]. It has a robust interpersonal character but is generally dominated by Firmicutes and Bacteroidetes, representing up to 90% of its composition and Actinobacteria, Proteobacteria, and Verrucomicrobia phyla [75].
However, even in stable chronological periods, the microbiota is subject to conditions capable of destabilizing homeostasis with the host. This process is called dysbiosis, a compositional and functional change in the microbiota that is driven by a set of factors that disturb the microbial ecosystem to a certain extent that exceeds its capacity for resistance and resilience. Several factors are associated with this phenomenon, including infections, diet, xenobiotics, genetics, familial transmission, circadian disruption, high-fat maternal diet, pregnancy, and physical injury [76].
An ecosystem in dysbiosis can damage the host immune system through various mechanisms that collectively stabilize the dysbiotic configuration. These mechanisms include modulation of inflammatory signaling by microbial metabolites, modulation of Toll-like receptor signaling (TLR), and degradation of IgA secreting agent (sIgA). The result is an intestinal epithelium more susceptible to pathobionts and disruption of the immune system, which decreases the protective capacity of the intestinal barrier, stimulating local or systemic inflammatory and immune-mediated processes [76,77,78]. Therefore, the genesis of several diseases had already been related to the microbiota’s dysbiotic configuration, including DM2, obesity, inflammatory bowel disease, cardiovascular diseases, autoimmune diseases, neurodegenerative diseases, NAFLD and its progressive form, non-alcoholic steatohepatitis (NASH) [79,80,81,82,83,84,85].
4. Microbiome, NAFLD, and NASH
4.1. Associations between the Gut Microbiome and NAFLD and NASH
The reciprocal interaction between the microbiome and the liver is established through the vascular route of the portal vein that takes to the liver gut-derived products and the liver feedback route of bile and antibody secretion to the intestine [15]. The liver is first exposed to gut-derived toxic factors, including bacteria, damaged metabolites, or bacterial products (LPS and bacteria DNA) [86].
Le roy et al. [87] demonstrated that when exposed to a high-fat diet, germ-free (GF) rats that received a transfer of gut microbiota from hyperglycaemic rats developed fasting hyperglycemia, hyperinsulinemia, and hepatic macrovesicular steatosis. In contrast, GF rats that received a transfer from normoglycemic rats remained normoglycaemic and without steatosis. Pyrosequencing of the 16S ribosomal RNA genes showed that hyperglycemic and normoglycemic rats had distinct gut microbiota regarding phylum, genus, and species. Henao-Mejia et al. [88] showed that sharing the microbiota through coprophagy from mice prone to developing NASH due to modifications in the inflammasome pathway in wild mice led to the development of steatosis and inflammation in the latter group.
Interestingly, some studies have observed that each stage of NAFLD corresponds to a pattern of gut microbiota [89]. A prospective study demonstrated that specific bacterial metagenomic signatures in the gut microbiome of NAFLD patients are a robust predictor of advanced fibrosis in humans. Some species were associated with NAFLD, with the abundance of bacterial species, such as Proteobacteria, Enterobacteria, Escherichia, and Bacteroides, being higher in NASH patients compared to matched healthy subjects [90].
There are several mechanisms by which the intestinal microbiota interfere in the progression of NAFLD and NASH. The increase in intestinal permeability, the translocation of dysbiotic bacteria, and the production of metabolites can be associated due to this dysbiotic state, and it is capable of generating disordered inflammatory responses which influence liver metabolism [91].
A new therapeutic attempt that has recently emerged is fecal microbiota transplantation. In this regard, Craven et al. [92] compared two groups: one received an autologous FMT, and the other received allogeneic FMT sourced from three lean, healthy individuals. At the end of the research, there was a significant decrease in the small intestinal permeability of the allogeneic group compared with the autologous group. Although the study did not show changes on biomarkers, it is known that increased intestinal permeability represents a central mechanism behind diseases such as inflammatory bowel disease, systemic inflammation, infection, MS, and NAFLD.
4.2. Microbiota-Derived Metabolites and Their Impact on NAFLD and NASH
Short-Chain Fatty Acids (SCFAs)
The short-chain fatty acids (SCFA) are the primary end products of fermentation of nondigestible carbohydrates (NDC) that become available to the gut microbiota. The main NDC are acetate, propionate, and butyrate [93,94]. Bacteroides are the main producers of propionate and acetate, while Firmicutes are the primary producers of butyrate [95]. Butyrate and propionate are well documented as gut inflammation relievers [96]. In rats, acetate and propionate supplementation decreased lipogenesis and fat accumulation, shielding them from high-fat (HF) diet-induced weight gain [97]. Svegliati-Baroni et al. [98] reported that the expression of the glucagon-like peptide-1 receptor (GLP-1r) is reduced in the hepatocytes of rats fed diet HF and patients with NASH, and that the activation of GLP-1r in the hepatocytes increased the oxidation of β-fatty acids and improved the insulin sensitivity. In addition, butyrate in particular is able to improve the function of tight junctions and stimulate mucin production, which helps maintain the integrity of the intestinal wall and prevent translocation of bacteria and its products, such as LPS, into the portal circulation [99,100].
Bile Acids
Bile acids are molecules produced in the liver from cholesterol and stored in the gallbladder. In addition to facilitating the absorption of lipids, they also play a role in glucose metabolism. The intestinal microbiota converts primary bile acids, including cholic acid (CA) and chenodeoxycholic acid (CDCA) in the distal small intestine and colon of humans into more than twenty different secondary bile acids, such as deoxycholic acid (DCA), lithocholic acid (LCA) and ursodeoxycholic acid (UDCA) [101,102]. Bile acids are indirectly involved in antimicrobial defense mediated by the farnesoid X receptor (FXR). Activation of this receptor reduces fatty acid and triglyceride synthesis in the liver by decreasing the expression of LXR and SREBP-1C [103]. FXR-deficient mice show reduced insulin sensitivity and decreased glucose tolerance [104]. In contrast, FXR activation by selective agonists suppresses bile acid and fatty acid production and increases glucose and insulin sensitivity in obese and diabetic mice. FXR activation also appears to attenuate primary biliary cirrhosis and NASH by reducing bile acid pool and liver fibrosis [105,106]. Bile acids are also closely related to another receptor, Takeda-G-protein-receptor-5 (TGR5). In the intestines, activation of TGR5 on L cells increases secretion of GLP-1, which binds to its receptor located on pancreatic beta cells, raising insulin secretion and reducing glucagon synthesis. The TGR5 is also able to modulate inflammatory processes. Its binding to the receptor reduces the release of pro-inflammatory cytokines by macrophages through the inhibition of NF-kB. Furthermore, in an experimental animal model, TGR5 knockout (TGR5-/-) mice have been shown to display accelerated LPS-induced inflammation in the liver and to suppress the inhibitory effect of TGR5 agonist on the expression of inflammatory mediators when compared with wild-type mice [107].
Choline and Trimethylamine
Choline is a constituent of the cells and mitochondrial membranes and the neurotransmitter acetylcholine. Choline-deficient diets have long been used to examine the mechanisms of fatty liver disease and its progression. They reproduce many of the phenotypes seen in humans with NAFLD, including an accumulation of triglycerides in the liver [108]. The phosphorylation of choline for the production of phospholipids and its oxidation as a methyl group donor are the main destinations of this nutrient [109]. Phosphatidylcholine is one of the most important choline metabolites. Its function is related to the packaging and export of triglycerides in very-low-density lipoprotein (VLDL) and to the solubilization of bile acids for excretion [110,111]. The lack of choline alters mitochondrial membranes, decreasing the concentration of phosphatidylethanolamine and phosphatidylcholine, which leads to a decrease in an action potential. This process consequently decreases ATP production and beta-oxidation, causing further hepatic steatosis [112,113]. For example, Arao et al. [114] used a methionine/choline-deficient diet to establish a NASH model and found that mitochondrial DNA content was decreased. The gut microbiome actively metabolizes choline, which may alter its bioavailability and potentially predispose one to choline deficiency [115]. The intestinal microbiota promotes the conversion of choline into trimethylamine, which, upon entering the circulation, will be converted into trimethylamine N-oxide in the liver [116]. Increased production of this substance results in a decrease in choline and consequently in the export of hepatic very-low-density lipoproteins and modulation of bile acid synthesis, which has detrimental effects on the liver, such as increased hepatic fat deposition and inflammatory and oxidative lesions and decreased glucose metabolism [89,117].
Ethanol
Ethanol is a microbial metabolite derived from saccharolytic fermentation. As late as 2000, Cope et al. [118] suggested that blood levels of ethanol were related to changes in the gut microbiota. Further, other studies have shown that dysbiosis in NASH patients involves ethanol-producing bacteria such as Escherchia coli, Bacteroides, Bifidobacterium, Clostridium [119,120]. One survey found a high increase in ethanol levels in NASH patients compared to healthy individuals or obese non-NASH patients [119]. Gut-bacteria-derived ethanol and its oxidized metabolite, acetaldehyde, are possibly involved in the progression of NAFLD through direct toxic effects on liver cells, through damage to the intestinal barrier generating increased portal endotoxemia, and through upregulation of nuclear factor-κB (NF-κB) signaling inflammatory pathways in peripheral cells [121,122]. Figure 3 shows the microbiota-derived metabolites and their impact on NAFLD and NASH.
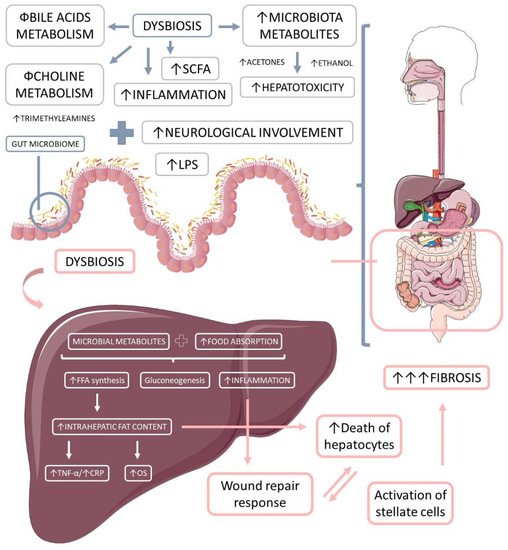
Figure 3. The microbiota-derived metabolites and their impact on liver inflammation, oxidative stress, and the development of liver diseases. Gut microbiome dysbiosis impairs the bile acids and choline metabolism, increases hepatotoxicity, and promotes inflammation. The microbiota-derived metabolites and the augmented food absorptions increase the intrahepatic production and accumulation of lipids, which causes increased inflammation and oxidative stress. Due to these events, the liver loses its capacity for wound repair response, and in addition to the augmented hepatocytes death and the augmented activation of stellate cells, liver massive fibrosis occurs. LPS: lipopolysaccharide; OS: oxidative stress, TNF-α: tumor necrosis factor-alpha; FFA: free fat acids; CRP: C reactive protein.
This entry is adapted from the peer-reviewed paper 10.3390/ijms23158805
This entry is offline, you can click here to edit this entry!