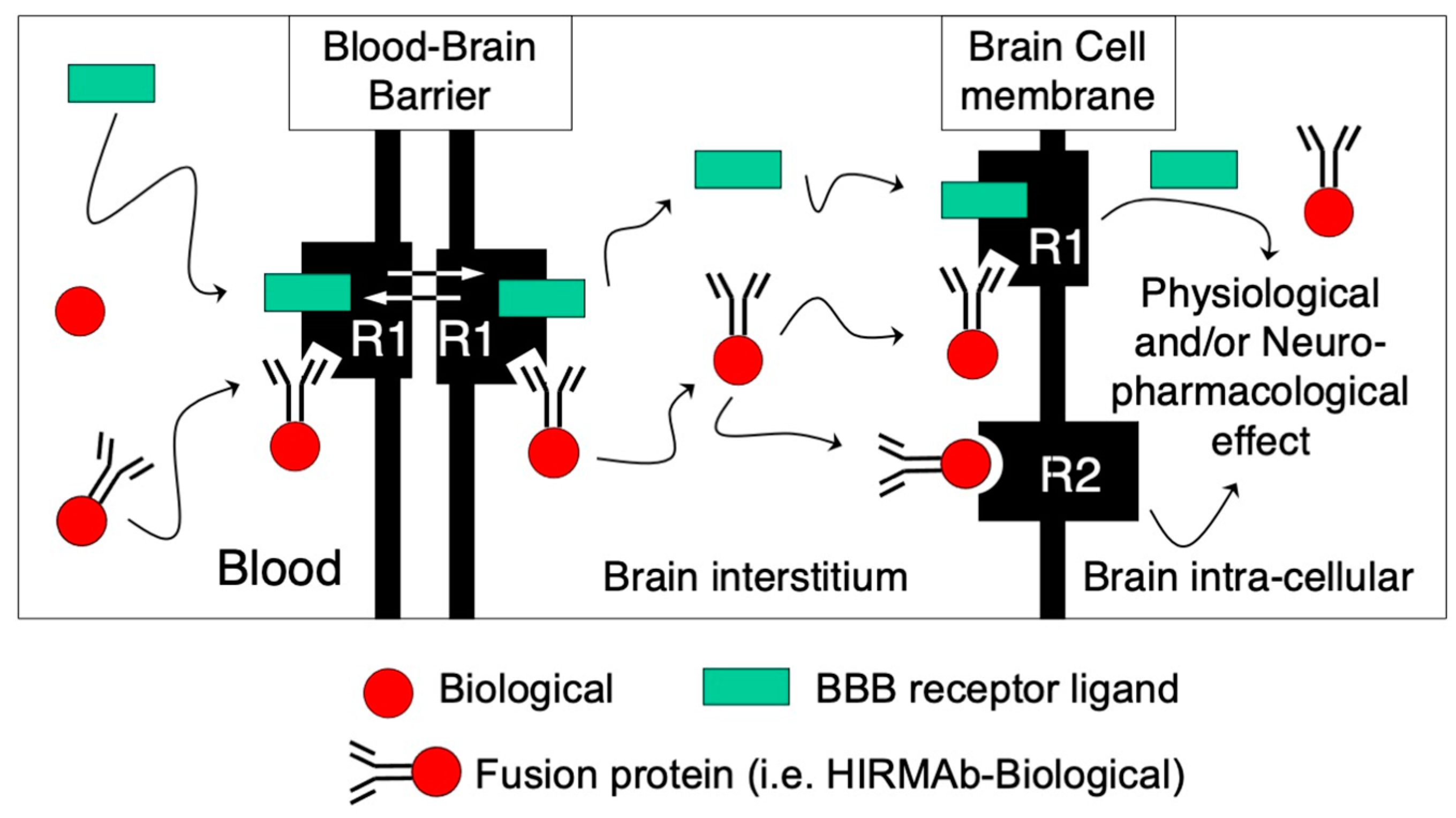
The treatment of neurological disorders with large-molecule biotherapeutics requires that the therapeutic drug be transported across the blood–brain barrier (BBB). Recombinant biotherapeutics, such as neurotrophins, enzymes, decoy receptors, and monoclonal antibodies (MAb), do not cross the BBB. These biotherapeutics can be re-engineered as brain-penetrating bifunctional IgG fusion proteins. These recombinant proteins comprise two domains, the transport domain and the therapeutic domain, respectively. The transport domain is an MAb that acts as a molecular Trojan horse by targeting a BBB-specific endogenous receptor that induces receptor-mediated transcytosis into the brain, such as the human insulin receptor (HIR) or the transferrin receptor (TfR).
- blood–brain barrier
- protein-based therapy
- monoclonal antibody
1. Introduction
| IgG Fusion Protein 1 | Therapeutic Domain | Indication | Reference |
|---|---|---|---|
| HIRMAb-IDUA (valanafusp alpha) | Iduronidase (IDUA) | Hurler syndrome (MPS I) | [22] |
| HIRMAb-IDS | Iduronate-2-sulfatase (IDS) | Hunter syndrome (MPS II) | [23] |
| TfRMAb-IDS (pabinafusp alfa) | Iduronate-2-sulfatase (IDS) | Hunter syndrome (MPS II) | [24] |
| HIRMAb-ASA | Arylsulfatase A (ASA) | Metachromatic leukodystrophy * | [25] |
| HIRMAb-SGSH | Sulfamidase (SGSH) | Sanfilippo A (MPSIIIA) * | [26] |
| HIRMAb-NAGLU | N-acetyl-alpha-D-glucosaminidase (NAGLU) | Sanfilippo B (MPSIIIB) * | [27] |
| HIRMAb-ASM | Acid shingomyelinase (ASM) | Niemann–Pick A/B * | [28] |
| HIRMAb-HEXA | Hexoaminidase A (HEXA) | Tay–Sachs * | [28] |
| HIRMAb-PPT1 | Palmitoyl-protein thioesterase (PPT1) | Batten Type 1 * | [28] |
| HIRMAb-GLB1 | β-galactosidase (GLB1) | GM1-gangliosidosis * | [28] |
| HIRMAb-Aβ bispecific antibody | Anti-Aβ amyloid single-chain Fv antibody (scFv) | Alzheimer’s * | [29] |
| HIRMAb-TNFR | Tumor necrosis factor decoy receptor (TNFR) | Parkinson’s, ALS, Alzheimer’s, and/or stroke * | [30] |
| HIRMAb-EPO | Erythropoietin (EPO) | Parkinson’s, Alzheimer’s, and/or Friedreich ataxia * | [31] |
| HIRMAb-GDNF | Glial-cell-derived neurotrophic factor (GDNF)) | Parkinson’s, stroke, and/or drug/EtOH addiction * | [32] |
| HIRMAb-BDNF | Brain-derived neurotrophic factor (BDNF) | Stroke, neural repair * | [33] |
| HIRMAb-Avidin | Any mono-biotinylated therapeutic | Various | [34] |
| IgG Fusion Protein 1 | Therapeutic Domain | Indication | Reference |
|---|---|---|---|
| TfRMAb-IDUA | Iduronidase (IDUA) | Hurler syndrome (MPS I) | [35] |
| TfRMAb-IDS | Iduronate-2-sulfatase (IDS) | Hunter syndrome (MPS II) | [36] |
| TfRMAb-SGSH | Sulfamidase (SGSH) | Sanfilippo A (MPSIIIA) * | [37] |
| TfRMAb-Aβ bispecific antibody | Anti-Aβ amyloid single-chain Fv antibody (scFv) | Alzheimer’s * | [38] |
| TfRMAb-TNFR | Tumor necrosis factor decoy receptor (TNFR) | Parkinson’s, Alzheimer’s, and/or stroke * | [39] |
| TfRMAb-EPO | Erythropoietin (EPO) | Parkinson’s, Alzheimer’s, and/or stroke * | [40] |
| TfRMAb-GDNF | Glial-cell-derived neurotrophic factor (GDNF)) | Parkinson’s, and/or stroke * | [41] |
| TfRMAb-Avidin | Any mono-biotinylated therapeutic | Various | [42] |
1 The transport domain of these mouse fusion proteins is a monoclonal antibody directed to the mouse BBB-transferrin receptor (TfRMAb). The therapeutic domain of the fusion protein and its experimental application are listed for the corresponding IgG fusion protein. * Indication has a primary CNS disease burden.
2. Genetic Engineering of IgG Fusion Proteins

This entry is adapted from the peer-reviewed paper 10.3390/pharmaceutics14071476
References
- Pardridge, W.M.; Boado, R.J.; Black, K.L.; Cancilla, P.A. Blood-brain barrier and new approaches to brain drug delivery. West. J. Med. 1992, 156, 281–286.
- Pardridge, W.M. The blood-brain barrier: Bottleneck in brain drug development. NeuroRx 2005, 2, 3–14.
- Abbott, N.J.; Patabendige, A.A.K.; Dolman, D.E.M.; Yusof, S.R.; Begley, D.J. Structure and function of the blood-brain barrier. Neurobiol. Dis. 2010, 37, 13–25.
- Pardridge, W.M. CSF, blood-brain barrier, and brain drug delivery. Expert Opin. Drug Deliv. 2016, 13, 963–975.
- Boado, R.J.; Pardridge, W.M. Molecular cloning of the bovine blood-brain barrier glucose transporter cDNA and demonstration of phylogenetic conservation of the 5′-untranslated region. Mol. Cell. Neurosci. 1990, 1, 224–232.
- Boado, R.J.; Li, J.Y.; Nagaya, M.; Zhang, C.; Pardridge, W.M. Selective expression of the large neutral amino acid trans-porter at the blood-brain barrier. Proc. Natl. Acad. Sci. USA 1999, 96, 12079–12084.
- Pardridge, W.M.; Eisenberg, J.; Yang, Y. Human blood-brain barrier insulin receptor. J. Neurochem. 1985, 44, 1771–1778.
- Fishman, J.B.; Rubin, J.B.; Handrahan, J.V.; Connor, J.R.; Fine, R.E. Receptor-mediated transcytosis of transferrin across the blood-brain barrier. J. Neurosci. Res. 1987, 18, 299–304.
- Boado, R.J.; Golden, P.L.; Levin, N.; Pardridge, W. Up-Regulation of Blood-Brain Barrier Short-Form Leptin Receptor Gene Products in Rats Fed a High Fat Diet. J. Neurochem. 2002, 71, 1761–1764.
- Duffy, K.R.; Pardridge, W.M.; Rosenfeld, R.G. Human blood-brain barrier insulin-like growth factor receptor. Metabolism 1988, 37, 136–140.
- Bickel, U.; Yoshikawa, T.; Landaw, E.M.; Faull, K.F.; Pardridge, W.M. Pharmacologic effects in vivo in brain by vector-mediated peptide drug delivery. Proc. Natl. Acad. Sci. USA 1993, 90, 2618–2622.
- Kang, Y.S.; Bickel, U.; Pardridge, W.M. Pharmacokinetics and saturable blood-brain barrier transport of biotin bound to a conjugate of avidin and a monoclonal antibody to the transferrin receptor. Drug Metab. Dispos. 1994, 22, 99–105.
- Wu, D.; Boado, R.J.; Pardridge, W.M. Pharmacokinetics and blood-brain barrier transport of -biotinylated phos-phorothioate oligodeoxynucleotide conjugated to a vector-mediated drug delivery system. J. Pharmacol. Exp. Ther. 1996, 276, 206–211.
- Wu, D.; Yang, J.; Pardridge, W.M. Drug targeting of a peptide radiopharmaceutical through the primate blood-brain barrier in vivo with a monoclonal antibody to the human insulin receptor. J. Clin. Investig. 1997, 100, 1804–1812.
- Xia, C.-F.; Zhang, Y.; Zhang, Y.; Boado, R.J.; Pardridge, W.M. Intravenous siRNA of Brain Cancer with Receptor Targeting and Avidin–Biotin Technology. Pharm. Res. 2007, 24, 2309–2316.
- Coloma, M.J.; Lee, H.J.; Kurihara, A.; Landaw, E.M.; Boado, R.J.; Morrison, S.L.; Pardridge, W.M. Transport across the primate blood-brain barrier of a genetically engineered chimeric monoclonal antibody to the human insulin receptor. Pharm. Res. 2000, 17, 266–274.
- Boado, R.J.; Zhang, Y.; Wang, Y.; Pardridge, W.M. Engineering and expression of a chimeric transferrin receptor mono-clonal antibody for blood-brain barrier delivery in the mouse. Biotechnol. Bioeng. 2009, 102, 1251–1258.
- Boado, R.J. A new generation of neurobiological drugs engineered to overcome the challenges of brain drug delivery. Drug News Perspect. 2008, 21, 489–503.
- Pardridge, W.M. Re-Engineering Biopharmaceuticals for Delivery to Brain with Molecular Trojan Horses. Bioconjug. Chem. 2008, 19, 1327–1338.
- Pardridge, W.M.; Boado, R.J. Reengineering Biopharmaceuticals for Targeted Delivery Across the Blood–Brain Barrier. Methods Enzymol. 2012, 503, 269–292.
- Pardridge, W.; Chou, T. Mathematical Models of Blood-Brain Barrier Transport of Monoclonal Antibodies Targeting the Transferrin Receptor and the Insulin Receptor. Pharmaceuticals 2021, 14, 535.
- Boado, R.J.; Zhang, Y.; Zhang, Y.; Xia, C.-F.; Wang, Y.; Pardridge, W.M. Genetic engineering of a lysosomal enzyme fusion protein for targeted delivery across the human blood-brain barrier. Biotechnol. Bioeng. 2007, 99, 475–484.
- Lu, J.Z.; Boado, R.J.; Hui, E.K.; Zhou, Q.H.; Pardridge, W.M. Expression in CHO cells and pharmacokinetics and brain uptake in the Rhesus monkey of an IgG-iduronate-2-sulfatase fusion protein. Biotechnol. Bioeng. 2011, 108, 1954–1964.
- Sonoda, H.; Morimoto, H.; Yoden, E.; Koshimura, Y.; Kinoshita, M.; Golovina, G.; Takagi, H.; Yamamoto, R.; Minami, K.; Mizoguchi, A.; et al. A Blood-Brain-Barrier-Penetrating Anti-human Transferrin Receptor Antibody Fusion Protein for Neuronopathic Mucopolysaccharidosis II. Mol. Ther. 2018, 26, 1366–1374.
- Boado, R.J.; Lu, J.Z.; Hui, E.K.-W.; Sumbria, R.K.; Pardridge, W.M. Pharmacokinetics and brain uptake in the rhesus monkey of a fusion protein of arylsulfatase a and a monoclonal antibody against the human insulin receptor. Biotechnol. Bioeng. 2012, 110, 1456–1465.
- Boado, R.J.; Lu, J.Z.; Hui, E.K.-W.; Pardridge, W.M. Insulin Receptor Antibody—Sulfamidase Fusion Protein Penetrates the Primate Blood–Brain Barrier and Reduces Glycosoaminoglycans in Sanfilippo Type A Cells. Mol. Pharm. 2014, 11, 2928–2934.
- Boado, R.J.; Lu, J.Z.; Hui, E.K.-W.; Lin, H.; Pardridge, W.M. Insulin Receptor Antibody−α-N-Acetylglucosaminidase Fusion Protein Penetrates the Primate Blood–Brain Barrier and Reduces Glycosoaminoglycans in Sanfilippo Type B Fibroblasts. Mol. Pharm. 2016, 13, 1385–1392.
- Boado, R.J.; Lu, J.Z.; Hui, E.K.; Lin, H.; Pardridge, W.M. Bi-functional IgG-lysosomal enzyme fusion proteins for brain drug delivery. Sci. Rep. 2019, 9, 18632.
- Boado, R.J.; Zhang, Y.; Zhang, Y.; Xia, C.F.; Pardridge, W.M. Fusion antibody for Alzheimer’s disease with bidirectional transport across the blood-brain barrier and Aβ fibril disaggregation. Bioconjug. Chem. 2007, 18, 447–455.
- Boado, R.J.; Hui, E.K.-W.; Lu, J.Z.; Zhou, Q.-H.; Pardridge, W.M. Selective targeting of a TNFR decoy receptor pharmaceutical to the primate brain as a receptor-specific IgG fusion protein. J. Biotechnol. 2010, 146, 84–91.
- Boado, R.J.; Hui, E.K.-W.; Lu, J.Z.; Pardridge, W.M. Drug Targeting of Erythropoietin Across the Primate Blood-Brain Barrier with an IgG Molecular Trojan Horse. J. Pharmacol. Exp. Ther. 2010, 333, 961–969.
- Boado, R.J.; Zhang, Y.; Zhang, Y.; Wang, Y.; Pardridge, W.M. GDNF fusion protein for targeted-drug delivery across the human blood–brain barrier. Biotechnol. Bioeng. 2007, 100, 387–396.
- Boado, R.J.; Zhang, Y.; Zhang, Y.; Pardridge, W.M. Genetic engineering, expression, and activity of a fusion protein of a human neurotrophin and a molecular Trojan horse for delivery across the human blood-brain barrier. Biotechnol. Bioeng. 2007, 97, 1376–1386.
- Boado, R.J.; Zhang, Y.; Zhang, Y.; Xia, C.F.; Wang, Y.; Pardridge, W.M. Genetic engineering, expression, and activity of a chimeric monoclonal antibody-avidin fusion protein for receptor-mediated delivery of biotinylated drugs in humans. Bioconjug. Chem. 2008, 19, 731–739.
- Boado, R.J.; Hui, E.K.-W.; Lu, J.Z.; Zhou, Q.-H.; Pardridge, W.M. Reversal of Lysosomal Storage in Brain of Adult MPS-I Mice with Intravenous Trojan Horse-Iduronidase Fusion Protein. Mol. Pharm. 2011, 8, 1342–1350.
- Zhou, Q.H.; Boado, R.J.; Lu, J.Z.; Hui, E.K.; Pardridge, W.M. Brain-penetrating IgG-iduronate 2-sulfatase fusion protein for the mouse. Drug Metab. Dispos. 2012, 40, 329–335.
- Boado, R.J.; Lu, J.Z.; Hui, E.K.-W.; Pardridge, W.M. Reduction in Brain Heparan Sulfate with Systemic Administration of an IgG Trojan Horse–Sulfamidase Fusion Protein in the Mucopolysaccharidosis Type IIIA Mouse. Mol. Pharm. 2017, 15, 602–608.
- Boado, R.J.; Zhou, Q.H.; Lu, J.Z.; Hui, E.K.; Pardridge, W.M. Pharmacokinetics and brain uptake of a genetically engi-neered bifunctional fusion antibody targeting the mouse transferrin receptor. Mol. Pharm. 2010, 7, 237–244.
- Zhou, Q.H.; Sumbria, R.; Hui, E.K.; Lu, J.Z.; Boado, R.J.; Pardridge, W.M. Neuroprotection with a brain-penetrating bio-logic tumor necrosis factor inhibitor. J. Pharmacol. Exp. Ther. 2011, 339, 618–623.
- Zhou, Q.-H.; Boado, R.J.; Lu, J.Z.; Hui, E.K.-W.; Pardridge, W.M. Re-Engineering Erythropoietin as an IgG Fusion Protein That Penetrates the Blood−Brain Barrier in the Mouse. Mol. Pharm. 2010, 7, 2148–2155.
- Zhou, Q.-H.; Boado, R.J.; Lu, J.Z.; Hui, E.K.-W.; Pardridge, W.M. Monoclonal Antibody-Glial-Derived Neurotrophic Factor Fusion Protein Penetrates the Blood-Brain Barrier in the Mouse. Drug Metab. Dispos. 2010, 38, 566–572.
- Zhou, Q.H.; Lu, J.Z.; Hui, E.K.; Boado, R.J.; Pardridge, W.M. Delivery of a peptide radiopharmaceutical to brain with an IgG-avidin fusion protein. Bioconjug. Chem. 2011, 22, 611–618.
- Boado, R.J.; Pardridge, W.M. Genetic engineering of IgG-glucuronidase fusion proteins. J. Drug Target. 2010, 18, 205–211.
- Li, J.Y.; Sugimura, K.; Boado, R.J.; Lee, H.J.; Zhang, C.; Duebel, S.; Pardridge, W.M. Genetically engineered brain drug delivery vectors: Cloning, expression and in vivo application of an anti-transferrin receptor single chain anti-body-streptavidin fusion gene and protein. Protein Eng. 1999, 12, 787–796.
- Pardridge, W.M.; Kang, Y.-S.; Buciak, J.L.; Yang, J. Human insulin receptor monoclonal antibody undergoes high affinity binding to human brain capillaries in vitro and rapid transcytosis through the blood-brain barrier in vivo in the primate. Pharm. Res. 1995, 12, 807–816.
- Uchida, Y.; Ohtsuki, S.; Katsukura, Y.; Ikeda, C.; Suzuki, T.; Kamiie, J.; Terasaki, T. Quantitative targeted absolute proteomics of human blood-brain barrier transporters and receptors. J. Neurochem. 2011, 117, 333–345.
- Hoshi, Y.; Uchida, Y.; Tachikawa, M.; Inoue, T.; Ohtsuki, S.; Terasaki, T. Quantitative atlas of blood-brain barrier trans-porters, receptors, and tight junction proteins in rats and common marmoset. J. Pharm. Sci. 2013, 102, 3343–3355.
- Boado, R.J.; Hui, E.K.-W.; Lu, J.Z.; Pardridge, W.M. AGT-181: Expression in CHO cells and pharmacokinetics, safety, and plasma iduronidase enzyme activity in Rhesus monkeys. J. Biotechnol. 2009, 144, 135–141.
- Ullman, J.C.; Arguello, A.; Getz, J.A.; Bhalla, A.; Mahon, C.S.; Wang, J.; Giese, T.; Bedard, C.; Kim, D.J.; Blumenfeld, J.R.; et al. Brain delivery and ac-tivity of a lysosomal enzyme using a blood-brain barrier transport vehicle in mice. Sci. Transl. Med. 2020, 12, 1163.
- Niewoehner, J.; Bohrmann, B.; Collin, L.; Urich, E.; Sade, H.; Maier, P.; Rueger, P.; Stracke, J.O.; Lau, W.; Tissot, A.C.; et al. Increased Brain Penetration and Potency of a Therapeutic Antibody Using a Monovalent Molecular Shuttle. Neuron 2014, 81, 49–60.
- Ng, P.P.; Dela Cruz, J.S.; Sorour, D.N.; Stinebaugh, J.M.; Shin, S.U.; Shin, D.S.; Morrison, S.L.; Penichet, M.L. An anti-transferrin receptor-avidin fusion protein exhibits both strong proapoptotic activity and the ability to deliver various molecules into cancer cells. Proc. Natl. Acad. Sci. USA 2002, 99, 10706–10711.
- Ng, P.P.; Helguera, G.; Daniels, T.R.; Lomas, S.Z.; Rodriguez, J.A.; Schiller, G.; Bonavida, B.; Morrison, S.L.; Penichet, M.L. Molecular events contributing to cell death in malignant human hematopoietic cells elicited by an IgG3-avidin fusion protein targeting the transferrin receptor. Blood 2006, 108, 2745–2754.
- Zhou, Q.-H.; Boado, R.J.; Hui, E.K.-W.; Lu, J.Z.; Pardridge, W.M. Chronic Dosing of Mice with a Transferrin Receptor Monoclonal Antibody-Glial-Derived Neurotrophic Factor Fusion Protein. Drug Metab. Dispos. 2011, 39, 1149–1154.
- Yamamoto, R.; Yoden, E.; Tanaka, N.; Kinoshita, M.; Imakiire, A.; Hirato, T.; Minami, K. Nonclinical safety evaluation of pabinafusp alfa, an anti-human transferrin receptor antibody and iduronate-2-sulfatase fusion protein, for the treatment of neuronopathic mucopolysaccharidosis type II. Mol. Genet. Metab. Rep. 2021, 27, 100758.
- Pardridge, W.M. Kinetics of Blood–Brain Barrier Transport of Monoclonal Antibodies Targeting the Insulin Receptor and the Transferrin Receptor. Pharmaceuticals 2021, 15, 3.
- Boado, R.J.; Hui, E.K.; Lu, J.Z.; Pardridge, W.M. IgG-enzyme fusion protein: Pharmacokinetics and anti-drug antibody response in Rhesus monkeys. Bioconjug. Chem. 2013, 24, 97–104.
- Sun, J.; Boado, R.J.; Pardridge, W.M.; Sumbria, R.K. Plasma pharmacokinetics of high-affinity transferrin receptor anti-body-erythropoietin fusion protein is a function of effector attenuation in mice. Mol. Pharm. 2019, 16, 3534–3543.