Pancreatic β-cells are critical in maintaining euglycemia by responding to blood glucose oscillations. β-cell dysfunction is a feature of diabetes mellitus which is characterized by abnormally high blood glucose levels. β-cells are responsible for the synthesis, storage and secretion of insulin, a hormone that is paramount for metabolic homeostasis. One hundred years after the successful isolation of insulin by Banting and collaborators, studies on insulin and β-cells have determined many of the factors needed for normal β-cell function, which is best defined by the appropriate expression of canonical β-cell factors involved in glucose-stimulated insulin secretion. One of these factors is the MAF bZIP transcription factor A (MafA). MafA, a large basic leucine zipper (bZIP) transcription factor (TF), derives its name from the viral oncogene v-maf, which was isolated from musculoaponeurotic fibrosarcoma in chicken
[1]. The seven members of the MAF family were subsequently identified by homology of their bZIP domain with v-maf and are divided into two subgroups according to molecular size: large MAFs (Maf, MafA, MafB and Nlr) and small MAFs (MafF, MafG and MafK), which lack a transactivation domain. These factors have roles in development and terminal differentiation across multiple tissues
[2][3][4], but have also been identified in disease pathogenesis
[5]. For instance, MafA was first identified as the trans-acting factor binding the RIPE3b/C1 element of the insulin promoter
[6][7][8] and has since been characterized as an important TF for β-cell function that is dysregulated in diabetes progression
[9][10]. These studies and others emphasized the critical role of MafA for β-cell function and suggested that elucidating mechanisms of MafA regulation would deepen the understanding of β-cell pathophysiology and potentially uncover therapeutic targets for the amelioration of β-cell dysfunction and diabetes progression.
2. MafA Regulates β-Cell Function
The importance of MafA for glucose homeostasis was shown in MafA-deficient mice, which displayed glucose intolerance after weaning due to impaired glucose-stimulated insulin secretion (GSIS) and age-dependent diabetes progression
[11][12]. MafA-deficient mice had decreased mRNA levels of
Ins1, Ins2, Pdx1, Neurod1 and
Slc2a2, suggesting that MafA is essential for the transcriptional identity of β-cells. MafA works synergistically with Neurod1 and Pdx1 to transactivate the insulin promoter, which is an effect that could not be fully replicated with MafB nor Maf
[13]. Along these lines, MafA promotes insulin transcription when overexpressed in rat islets
[14] and when ectopically expressed in chick embryonic endoderm
[15] or in other non-β-cell lines of endodermal origin
[16][17].
Several studies suggest that MafA is dispensable for β-cell development but plays an essential role in β-cell maturation and glucose responsiveness of adult β-cells. In rodents,
Mafa expression is low at birth, increasing during the postnatal period
[18][19]. Interestingly, premature expression of MafA in pancreatic endocrine progenitors prevents differentiation into hormone-expressing cells
[20][21], suggesting that the induction of MafA expression in β-cells is time sensitive. Consistently, in mice with whole pancreas MafA knockout
(Mafa∆panc), the effects of MafA loss are not apparent until three weeks of age, when the KO mice showed lower β-cell mass, decreased insulin expression, and impaired glucose tolerance compared to controls
[22]. Similarly, at twelve weeks of age in whole-body MafA knockout mice, there is a reduced β-cell:α-cell ratio, decreased islet insulin content and β-cell dedifferentiation into MafB-expressing, progenitor-like cells
[12]. These results may recapitulate some aspects of normal islet development. MafB is actually the predominant large MAF protein expressed during pancreatic development in both α- and β-cells
[4]. MafB can upregulate
Ins1 and
Ins2 transcription
[8], but in adult mice, MafB is mainly expressed in α-cells and upregulates
Gcg gene expression
[4][23]. The decrease in β-cell MafB expression after birth is at least partly due to methylation, specifically by DNA methyltransferase 3a (Dnmt3a) which binds and represses at a region −1032 to −838 upstream of the
Mafb transcriptional start site
[24].
Mafb∆panc mice have higher blood glucose levels at P1, but two weeks after birth, blood glucose levels normalize. Likewise, at E15.5,
Mafb∆panc mice have a lower number of insulin
+ and glucagon
+ cells compared to controls, but two weeks after birth, cell numbers became roughly the same
[25]. MafB does seem to have a role in enhancing postnatal β-cell function under metabolic stress (pregnancy, high-fat diet)
[24][26], but MafA remains critical for proper β-cell function and glucose homeostasis in mature β-cells, with forced MafB expression unable to compensate for MafA loss in adult mice
[24].
Reduced MafA expression is associated with diabetes progression in mice and human patients. In islets isolated from diabetic
db/db mice, MafA is decreased due to hyperglycemia-associated oxidative stress and c-Jun activity
[9]. Similarly, islets from subjects with type 2 diabetes (T2D) display a marked decrease in
MAFA mRNA levels and protein expression
[10][27][28]. Several studies have shown possible mechanisms for this phenomenon. In β-cells from subjects with T2D
[29] and in in vitro studies mimicking oxidative stress
[10], MafA was found primarily in the cytoplasm rather than properly localized in the nucleus. However, when MafA levels were “rescued” in
db/db mice, GSIS and β-cell mass improved, suggesting that preserving MafA expression in β-cells can still mitigate diabetes progression
[30]. scRNA-seq analysis of human β-cells supports this concept as well, wherein metabolically inflexible β-cells show lower MAFA activity as compared to healthy β-cells
[31]. This finding is thought to align with previous patterns of T2D-induced oxidative stress, in which case MafA activity is high under healthy islet conditions but can also increase as a protective mechanism against acute oxidative stress
[32]. In a different scRNA-seq analysis of human islets, β-cells co-expressing
MAFA/MAFB had increased expression of genes related to β-cell identity, glucose metabolism and exocytosis compared to β-cells that only expressed one or neither TF
[33]. Since MAFA and MAFB are capable of forming heterodimers
[8][34], it is possible that the co-expression of both can enhance the β-cell maturity transcriptional program. These and other studies have documented the functional and transcriptional heterogeneity of β-cells
[31][33][35].
3. MafA Target Genes
MafA transcriptional activity is necessary for proper postnatal β-cell function. MafA was first identified as a TF of insulin that binds to the C1 element of the insulin gene promoter. MAF proteins, including MafA, bind to the MAF-recognition elements (MAREs), through a DNA consensus sequence TGCTGAC(G)TCAGCA
[36]. However, in β-cells, the MARE sequence within the C1 element deviates slightly from the consensus sequence. The MARE sequence in C1 element of the rat and mouse
Ins2 promoter is TGCAGCTTCAGCC whereas the equivalent in the human
INS promoter is TGCAGCCTCAGCC (the underlined nucleotides indicate deviations from the MARE consensus sequence)
[7][16][37]. Interestingly, in addition to the C1 element, three other MARE sites (MARE1, MARE2 and MARE3) were also identified in the rat
Ins2 and human
INS promoter. MafA can bind to MARE2 and MARE3 in the rat
Ins2 promoter while MafA can bind to MARE1 in the human
INS promoter
[16].
Although MafA can activate the insulin promoter alone, the addition of Neurod1 and Pdx1, which bind the E box and A box of the insulin promoter, respectively, synergistically promote insulin transcription
[13]. Consistently, mice with homozygous CRISPR-Cas9-introduced mutations in the C1 box of the
Ins1 and
Ins2 gene promoters are glucose intolerant. These C1 box mutations also prevented MafA from activating the promoter in luciferase assays, further supporting the importance of MafA in upregulating the promoter of its target genes
[38]. Human mutations characterized by a deletion of the C1 and E1 elements of the
INS promoter have been shown to cause permanent neonatal diabetes, which may be attributed to the disruption of MAFA and NEUROD1 binding sites
[39][40]. Other factors are also involved in this regulation. For example, Gli-similar 3 (Glis3), a Krüppel-like zinc finger transcription factor, activates insulin transcription by recruiting histone acetyltransferase Creb-binding protein (CBP) and by acting as a scaffold for the MafA, NeuroD1 and Pdx1 complex
[41].
Beyond insulin, MafA increases the expression of a wide range of genes involved in β-cell function, including genes implicated in glucose sensing, insulin processing, Ca2+ influx and oxidative phosphorylation. Many studies have identified a plethora of MafA target genes, all having important roles in proper β-cell function, further supporting MafA’s critical role in β-cells. While some studies have found direct MafA target genes (i.e., MafA directly binds to the promoter of the target genes), other studies describe genes that are differentially expressed when MafA protein is overexpressed or silenced, without showing direct MafA binding to the target gene promoters (i.e., MafA knockout or overexpression causes a decrease or increase in the target gene expression, respectively). Some of these MafA-target genes are summarized in Table 1. The lack of promoter binding evidence does not necessarily mean MafA cannot bind to the promoter of these genes, but rather that MafA may regulate these genes through intermediaries rather than direct promoter binding.
Table 1. MafA target genes. (*) denotes evidence that MafA directly binds target gene promoter.
|
Gene Symbol
|
Gene Name
|
Gene Function
|
Reference
|
|
Cacng4 *
|
Voltage-dependent calcium channel gamma-4 subunit
|
Enhances L-type Ca2+-mediated Ca2+ entry into β-cell
|
[42]
|
|
Chrnb4 *
|
Cholinergic receptor nicotinic beta 4
|
Subunit of the nicotinic acetylcholine receptor
|
[43]
|
|
Cox6a2 *
|
Cytochrome C oxidase subunit 6A2
|
One out of 13 subunits of cytochrome C oxidase complex (Complex IV), the last enzyme in the electron transport chain
|
[44]
|
|
G6pc2 *
|
Glucose-6-phosphate catalytic subunit related protein
|
Islet-specific enzyme that hydrolyzes glucose-6-phosphate, limits basal insulin secretion
|
[45]
|
|
Gck
|
Glucokinase
|
Phosphorylates glucose to glucose-6-phosphate in pancreatic islets and hepatocytes. Considered the β-cell glucose sensor
|
[46]
|
|
Glp1r
|
Glucagon-like peptide 1 receptor
|
Receptor for Glucagon-like peptide 1 (Glp1), a stimulator of insulin secretion
|
[46]
|
|
Ins1 *
|
Insulin I
|
One of two insulin genes in mouse, on chromosome 19
|
[11]
|
|
Ins2 *
|
Insulin II
|
One of two insulin genes in mouse, on chromosome 7
|
[11]
|
|
Maob *
|
Monoamine oxidase B
|
Metabolizes monoamine neurotransmitters, specifically benzylamine, dopamine and phenylethylamine
|
[47]
|
|
Neurod1
|
Neurogenic differentiation 1
|
Transactivator of genes important for β-cell maturation and function, including insulin
|
[46]
|
|
Nkx6.1
|
NK6 homeobox1
|
TF involved in β-cell development and regulation of genes involved in mature β-cell function
|
[46][48]
|
|
Pcsk1
|
Proprotein convertase subtilisin/kexin type 1
|
Proprotein convertase, which processes proinsulin in β-cells
|
[46]
|
|
Pcx
|
Pyruvate carboxylase
|
Catalyzes the conversion of pyruvate to oxaloacetate
|
[46]
|
|
Pdx1 *
|
Pancreatic and duodenal homeobox 1
|
TF important pancreas development and for mature β-cell function
|
[46][49]
|
|
PPP1R1A
|
Protein phosphatase 1, regulatory inhibitor subunit 1A
|
Regulates cAMP/PKA signaling pathway
Promotes Glp1-induced GSIS
|
[50]
|
|
Prlr
|
Prolactin Receptor
|
Involved in increasing β-cell mass during pregnancy
|
[51]
|
|
Slc2a2 *
|
Solute Carrier Family 2 Member 2
|
Glucose transporter 2, transmembrane glucose transporter with a high Km for glucose
|
[46][52]
|
|
Slc80a8
|
Solute carrier family 30 member 8
|
Zinc transporter on insulin granules in β-cells
|
[19][43]
|
4. Regulation of MafA Transcription
The first level of MafA regulation is through
Mafa gene expression. The
Mafa promoter consists of six regions (R1-R6), which contain highly conserved sequences 25 kb upstream the 5′ MafA untranslated region (UTR). R1 spans from −9389 to −9194, R2 spans from −8420 to −8293, R3 spans from −8118 to −7750, R4 spans from −6622 to −6441, R5 spans from −6217 to −6031 and R6 spans from −250 to +56. Of these, R3 has been shown to be most necessary, but not sufficient, for transcriptional initiation mediated by direct TF binding
[53][54]. Many transcription factors bind R3 on the
Mafa promoter, including Pax6, Nkx6.1, Nkx2.2, Pdx1, Hnf1a, Foxa2 and Isl1. Changes in these transcription factors directly correlate with
Mafa expression. However, there are some exceptions. Neurod1, another important insulin transcription factor, can bind to R3 of the
Mafa promoter, but
Neurod1−/− mice do not have detectable changes in MafA staining
[55]. However, in a more recent study,
Neurod1∆endo mice had lower Mafa mRNA levels, suggesting that Neurod1 may regulate
Mafa transcription
[56]. Additionally, Pax4 is a negative regulator of
Mafa. Pax4 overexpression limits R3-mediated promoter activity, but there is limited evidence of direct Pax4 binding to R3 via ChIP, possibly due to Pax4 regulating
Mafa in an indirect manner
[57].
Other TFs bind elsewhere in the MafA promoter or regulate
Mafa transcription through other mechanisms, as summarized in
Table 2. For instance, Onecut1, a TF involved in pancreatic endocrine development that is upregulated in adult
db/db mice, acts as a negative regulator of
Mafa transcription by preventing Foxa2 from interacting with the
Mafa promoter, highlighting the importance of the transcriptional regulation of
Mafa in metabolic disease states
[58].
Table 2. Known transcriptional regulators of Mafa gene expression.
|
Protein Symbol
|
Protein Name
|
Mafa Transcription Mechanism
|
Reference
|
|
Pax6
|
Paired box protein Pax-6
|
Binds R1, R3 and R6 of the Mafa promoter
|
[55]
|
|
Nkx6.1
|
NK6 homeobox 1
|
Binds R3 of the Mafa promoter
|
[55]
|
|
Nkx2.2
|
NK2 homeobox 2
|
Binds R3 of the Mafa promoter
|
[54]
|
|
Pdx1
|
Pancreatic and duodenal homeobox 1
|
Binds R3 and R6 of the Mafa promoter
|
[54][55]
|
|
Hnf1a
|
Hepatocyte nuclear factor 1-alpha
|
Binds R3 of the Mafa promoter
|
[53]
|
|
Foxa2
|
Forkhead box A2
|
Binds R3 of the Mafa promoter
|
[54]
|
|
Isl1
|
Insulin gene enhancer protein ISL-1
|
Binds R3 of the Mafa promoter
|
[59]
|
|
Neurod1
|
Neurogenic differentiation 1
|
Binds R3 of the Mafa promoter
|
[54]
|
|
Pax4
|
Paired box protein Pax-4
|
Negative regulator of Mafa, potentially by interfering other factors from binding R3 of the Mafa promoter
|
[57]
|
|
Mafb
|
Transcription factor MafB
|
Binds R3 of the Mafa promoter
|
[55][60]
|
|
Onecut1
|
One cut domain, family member 1
|
Prevents Foxa2 from binding to the Mafa promoter
|
[58]
|
|
Foxo1
|
Forkhead box O1
|
Binds to the forkhead element of the Mafa promoter
|
[32]
|
|
Thra
|
Thyroid hormone receptor alpha
|
Binds to Thyroid hormone response element (TRE), which are located from −1927 to −1946 and from +647 to +659 (named Site 2 and Site 3, respectively)
|
[61]
|
|
CREB
|
cAMP responsive element binding protein
|
Constitutively binds to the cAMP response element (CRE), spanning from −1342 to −1346, of the Mafa promoter
|
[62]
|
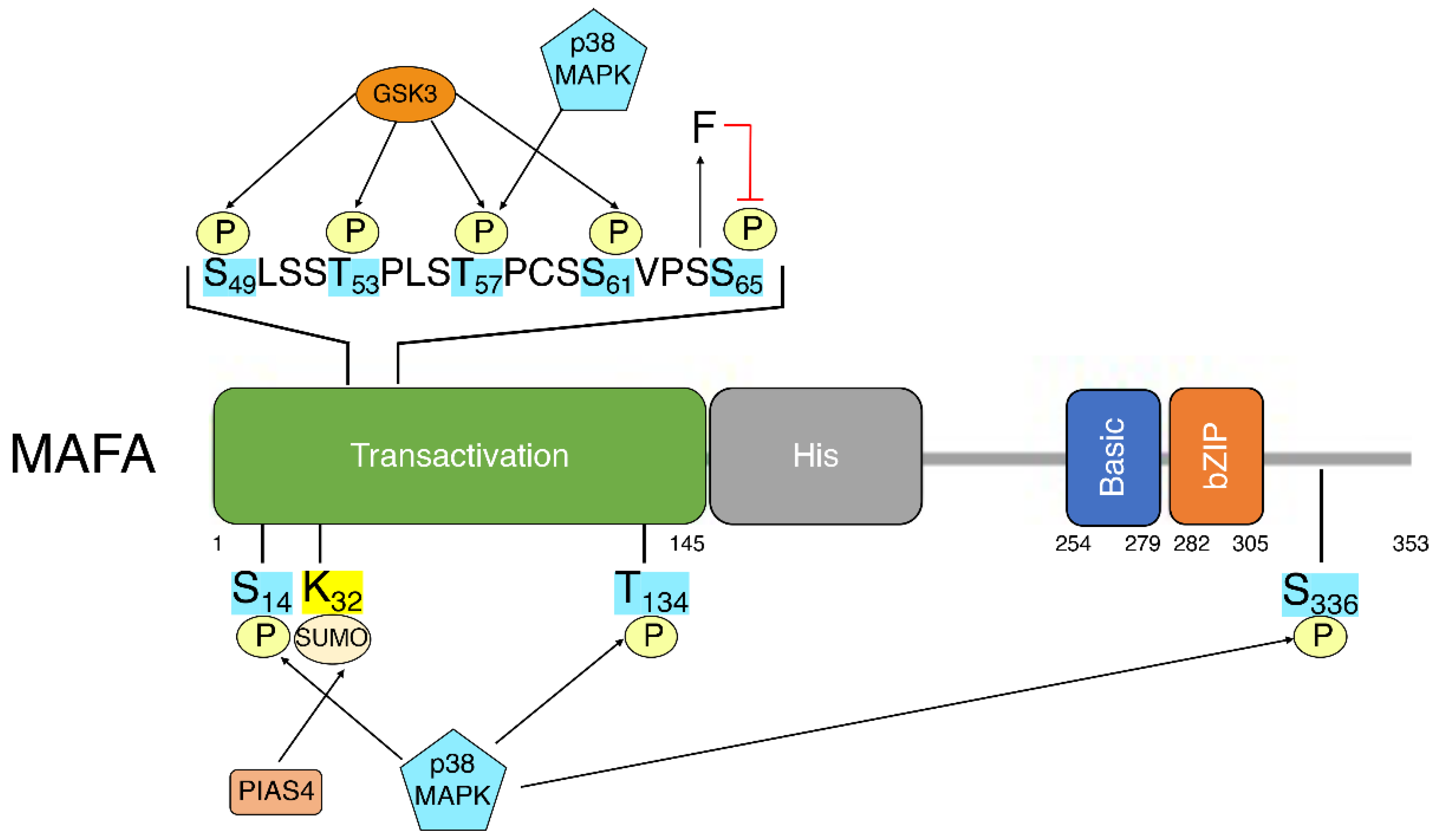
5. MafA Post-Translational Modifications
The previously discussed studies indicate the importance and potential mechanisms of
Mafa transcriptional regulation. Post-translational modifications (PTMs) add a second layer of regulation, allowing for dynamic changes in MafA activity or stability in response to changes in β-cell environment. While most documented PTMs are found in the N-terminal transactivation domain of MafA
[63], other regions of MafA, such as the extended homology region, the basic domain and the leucine-zipper, affect DNA recognition and binding and are also likely affected by the addition of PTMs
[5] (
Figure 1).
Figure 1. Linear schematic of MAFA depicting MAFA domains and known post-translationally modified residues. Transactivation: transactivation domain. His: histamine-rich region. Basic: basic domain. bZIP: Leucine zipper.