Accumulating evidence indicates that two major proteins are responsible for the structural coherence of bounding cardiomyocytes. These biomolecules are known as myocardial fibrillar collagen type I (COL1) and type III (COL3). In addition, fibronectin, laminin, fibrillin, elastin, glycoproteins, and proteoglycans take part in the formation of cardiac extracellular matrix (ECM). In physiological conditions, collagen synthesis and degradation in human cardiac ECM are well-regulated processes, but they can be impaired in certain cardiovascular diseases, such as heart failure (HF). Myocardial remodeling is part of the central mechanism of HF and involves cardiomyocyte injury and cardiac fibrosis due to increased fibrillar collagen accumulation.
- extracellular matrix
- collagen type I and III derived peptides
- heart failure
1. Introduction
2. Type I and Type III Collagen Characteristics
3. Cardiac Extracellular Matrix: Structure and Function
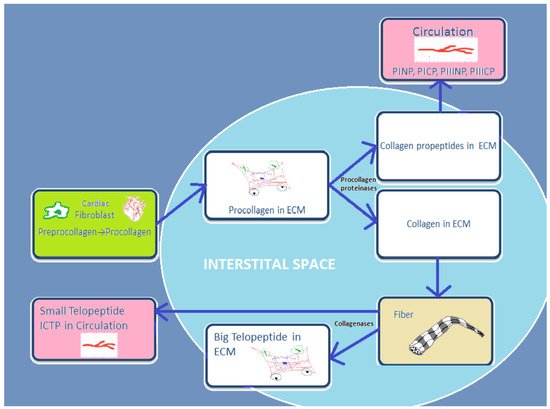
4. General Concepts of Abnormal Cardiac Extracellular Matrix Changes in Heart Failure
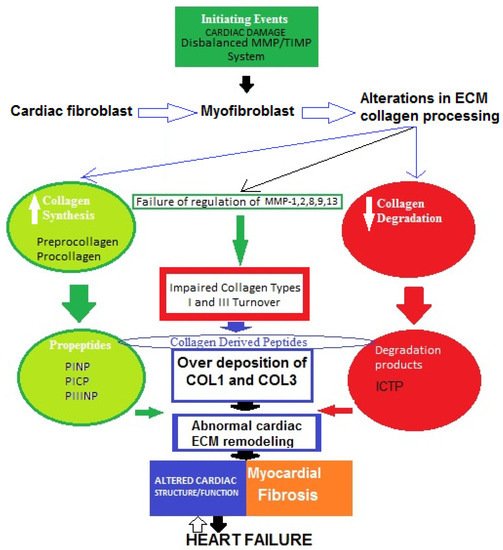
5. Basic Underlying Mechanisms of Myocardial Fibrosis in Heart Failure: Role of Impaired Type I and III Collagen Turnover
This entry is adapted from the peer-reviewed paper 10.3390/metabo12040297