Reticulocyte maturation begins after enucleation in the bone marrow. In rats, reticulocytes reside in the bone marrow from 6.5–17 hrs depending on the blood demand. These reticulocytes are termed as R1 and are characterized for their multi-lobular shape and their motility. The final stages of maturation occur during circulation where macrophages residing in the spleen may facilitate the process. These reticulocytes in circulation are termed as R2, are non-motile and have a “deep-dish” shape”. As part of their maturation, reticulocytes need to remove or degrade residual organelles and RNA. In addition, the reticulocyte must reduce its surface area and volume. On average, labelled baboon reticulocytes showed a reduction of 20% of their surface area and 15% of their volume after the first 24 h in circulation; at this point they showed a similar size distribution to that of mature RBC.
1. Protein Removal through Exosome Release
Multiple mechanisms have been shown to contribute to the process of reticulocyte maturation. Johnstone, Bianchini and Teng [
85] and Pan and Johnstone [
86], described the externalization of TfR in the form of vesicles during the maturation of sheep reticulocytes. They showed that this was achieved by a process of endocytosis of TfR from the surface of the plasma membrane into the cytoplasm. These newly formed endosomes fused together to form larger vesicle structures. Budding at the internal surface of the large vesicles resulted in the formation of multivesicular bodies (MVB) ranging from 0.5–1 μm in diameter, with intra-luminal vesicles (ILV) of ~50 nm in diameter [
87] The MVB would fuse with the plasma membrane releasing the internal vesicles termed exosomes (
Figure 4).
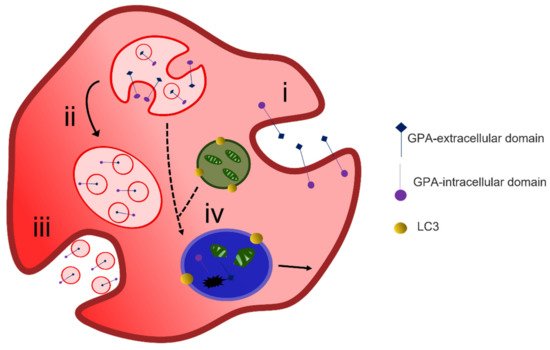
Figure 4. Exocytosis-mediated reticulocyte maturation. During reticulocyte maturation unwanted membrane proteins are endocytosed (i) and sorted into multivesicular bodies (ii) that are released as exosomes via the fusion of MVB membrane with the cell plasma membrane (iii). LC3-positive autophagosomes containing degraded organelles may also fuse with MVB and lysosomes (iv) and form autophagolysosomal vacuole where cargo degradation occurs or alternatively exits the cell via membrane budding.
Characterization of the exosomes released has shown that the removal of obsolete proteins is a selective process. Compartmentalization of exosome-derived proteins from cord blood reticulocytes showed that the main protein fractions were from the cytosol 15.9%, the plasma membrane 14.9% and the nucleus 13.5%. In equal parts (~5%) proteins were derived from the cytoskeleton, lysosomal, endosomal and Golgi compartments. Lower protein numbers were from the ER and the mitochondria [
88].
The sorting of membrane and cytosolic proteins into exosomes and not into the lysosomal pathway has been attributed to parallel mechanisms. The first one involves the binding of the cytosolic proteins Alix and heat shock cognate 70 to ubiquitinated proteins forming a link with the endosomal sorting for complex transport machinery (ESCRT, I and II). ESCRT III then promotes the formation of ILVs by assisting in pit formation and budding off the membrane [
89,
90]. Budding of exosomes from the intraluminal membrane of MVB has also been attributed to an ESCRT-independent system where lipid-raft micro-domains are formed in areas with ceramide enrichment [
91].
A second mechanism is also initiated by aggregation of receptors which has been shown to prevent re-joining the recycling pathway and the clusters may act as signals for exosome processing [
81]. Aggregation has also been promoted by the addition of exogenous lectins. Galectin-5 is removed from the surface during maturation of rat reticulocytes; this lectin is shown to enter the endosomal compartment from the cytosol and associate with the ILV surface. Lamp2, an endo-lysosomal protein which is also lost during reticulocyte maturation, has a binding motif for Galectin 5. Galectin-5 has also been shown to play a role in the sorting of glycoproteins into exosomes [
92]. Lipidomic studies on exosomes have suggested that all these pathways occur sequentially with the involvement of ceramides within lipid rafts featuring at the later stages of maturation [
89].
2. Alternative Methods of Reticulocyte Maturation
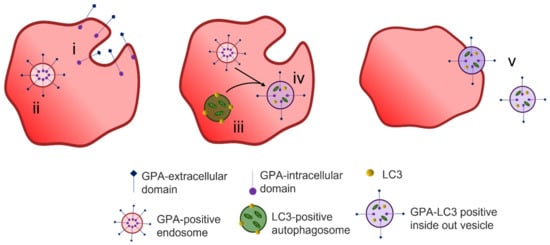
The process of exosome release to remove unwanted proteins does not fully explain the reduction in plasma membrane surface area during maturation of a reticulocyte to an erythrocyte. A two-stage alternative reticulocyte maturation mechanism has been proposed. In the bone marrow, the formation of an R2 reticulocyte from an R1 reticulocyte follows the removal of redundant proteins through the endosome–exosome pathway. For the final maturation, the R2 must remove excess plasma membrane. The plasma membrane is taken into the cytosol by endocytosis, and the GPA-positive containing endosome fuses with an autophagosome which is positive for the autophagosomal marker LC3; together they form an inside-out vesicle that is extruded through the plasma membrane at an area of weakened cytoskeleton [
93,
94] (
Figure 5). These inside-out vesicles are phosphatidylserine positive and stain with markers for organelles such as calreticulin for the ER, giantin for the Golgi and MitoTracker for the mitochondria [
95]. Their extrusion through the plasma membrane is thought to be driven by non-muscle myosin II [
96].
Figure 5. Reticulocyte maturation by expulsion of inside out hybrid vesicles. Endocytosis of superfluous membrane proteins occurs at the plasma membrane (i) during reticulocyte maturation forming GPA-positive endosomes (ii). LC3-positive autophagosomes are formed from isolated membranes that engulf cytoplasmic content including organelles (iii). The GPA-positive endosomes fuse with LC3-positive autophagosomes forming hybrid LC3-GPA positive inside out vesicles (iv) that are extruded during passage through the spleen (v). Figure modified from Mankelow et al. [
94].
Another model for membrane loss focusing on the role of membrane rafts for the latter stage of reticulocyte maturation has been described [
97]. Membrane rafts are known to be rich in cholesterol and sphingolipids. During early reticulocyte maturation these lipid rafts are gradually lost; however, differential expression of some membrane raft markers is observed in the mature erythrocyte membrane. This is suggested to be due to the presence of different type of lipid rafts. Areas of the membrane associated with flotillin-rich lipid rafts bulge and are removed by macrophages during circulation, resulting in removal of plasma membrane. Areas containing stomatin-rich lipid rafts are maintained within the membrane and allow anchoring to the cytoskeleton, which contributes to the formation of the final biconcave shape of a mature erythrocyte [
98].
3. Organelle Clearance
Autophagy is believed to be the system by which organelle clearance occurs. Autophagocytosis begins with the formation of a C-shaped membrane known as a phagophore that originates in the endoplasmic reticulum (ER), the mitochondria or the Golgi [
99]. During reticulocyte maturation, phagophore assembly is regulated by the autophagy-related proteins (Atg), in particular, Atg5 and Atg7. Although Atg5/Atg7-independent autophagy process has been shown to be involved for mitochondrial and ribosomal clearance, Ulk1 (Atg1)-deficient mice appear to retain mitochondria compared to wild type and Atg5-deficient mice [
100]. This Ulk1-dependent pathway appears to rely on regulatory Ras-associated binding proteins (Rab) that are involved in endosome fusion, transport, degradation and recycling [
101].
The role of the phagophore is to surround organelles due for elimination and enclose them in a double-membrane vesicle known as an autophagosome [
102]. The method of the autophagosome clearance remains controversial. In the canonical pathway the autophagosome fuses with lysosomes for cargo degradation by the action of hydrolytic enzymes. However, in erythroid cells, there is evidence of removal of the lysosomal compartment by disappearance of LAMP2 by exocytosis [
92]. As previously mentioned, a method of organelle elimination, combined with membrane reduction, was proposed to occur by the formation of large inside out hybrid vesicles that are extruded from the membrane to be removed by splenic macrophages [
95]. Alternatively, the interaction of autophagosomes with MVB has been proposed as a model for organelle clearance. In K562 cells, endocytic vesicles label with the autophagosome marker LC3 [
103,
104] and electron microscopy of rat reticulocytes shows that they contain TfR positive internal vesicles that have membrane continuity with autophagic vacuoles [
105].
This entry is adapted from the peer-reviewed paper 10.3390/membranes12030311