Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is an old version of this entry, which may differ significantly from the current revision.
Subjects:
Biochemistry & Molecular Biology
Mutualistic symbiosis refers to the symbiotic relationship between individuals of different species in which both individuals benefit from the association. S100A10, a member of the S100 family of Ca2+-binding proteins, exists as a tight dimer and binds two annexin A2 molecules. This association forms the annexin A2/S100A10 complex known as AIIt, and modifies the distinct functions of both proteins. Annexin A2 is a Ca2+-binding protein that binds F-actin, phospholipid, RNA, and specific polysaccharides such as heparin.
- S100A10
- annexin A2
- plasminogen
- plasmin
- symbiosis
1. Introduction
The nomenclature for the S100 family of Ca2+-binding proteins is derived from observing that these proteins are soluble in a saturated ammonium sulfate solution at a neutral pH. The S100 family of proteins consists of about twenty 10–12 kDa, Ca2+-binding proteins that are expressed only in vertebrates and consist of S100A1–S100A18, S100B, S100G, S100P, S100Z, as well as members with an N-terminal S100 domain fused to additional C-terminal sequences, such as repetin [1], trichohyalin and filaggrin [2,3,4]. Of the twenty-five human S100 genes, 19 are located within chromosome 1q21. Other members (S100A11, S100B, S100G, S100P, and S100Z) map to different chromosome regions [5]. Each S100 protein has a unique expression and distribution profile amongst different tissues and cell types. The S100 proteins exist as anti-parallel homo- and heterodimers, with each monomer consisting of two helix-loop-helix EF-hand motif types, Ca2+-binding domains (EF-1 and EF-2) connected by a hinge region. The nomenclature for the EF-hand domain originally comes from the EF-hand motif first identified in the Ca2+-binding protein, parvalbumin, which is composed of two alpha-helices, “E” and “F” connected by an intermediate loop of 12 residues binding Ca2+ [6]. Specifically, the S100 proteins possess an S100-specific EF-hand at the N-terminus and a canonical EF-hand at the C-terminus. In all S100 proteins except S100A10, the EF-hands bind Ca2+. These EF-hands are not identical; the carboxyl-terminal EF-hand Ca2+-binding loop contains the classical 12-amino-acid long Ca2+-binding motif common to all EF-hand Ca2+-binding proteins, such as parvalbumin and calmodulin. In this motif, Ca2+-binding occurs via acidic side chains that comprise the sequence DXDGDGTIXXXE. However, the N-terminal EF-hand motif is a 14-amino-acid long Ca2+-binding loop that is characteristic of all S100 proteins and is referred to as the S100-specific or pseudo-EF domain. This motif binds Ca2+ via backbone carbonyl groups and one carboxylate side group contributed by glutamic acid (reviewed in [2,7,8]). All S100 proteins bind calcium, except S100A10, which has lost its ability to bind calcium due to substitutions in its calcium-binding loop but retains the “active” conformation of a calcium-bound S100 protein [9]. S100A10 lost its ability to bind calcium due to substitutions in its calcium-binding loop but retains the activated structure of a calcium-bound S100 protein [10] (Figure 1).
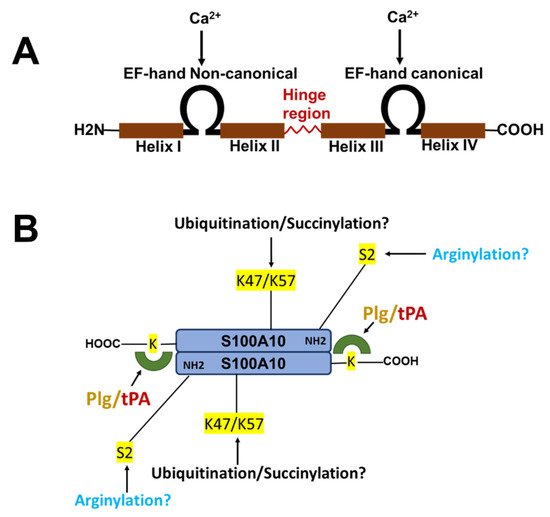
Figure 1. Structure of S100 proteins. (A) The structure of a typical S100 protein. (B) The structure and key regulatory sites of S100A10. Shown are the sites for ubiquitylation, arginylation, and succinylation.
The S100 proteins S100A1, S100A4, S100A6, S100A10, S100A11, S100A12, and S100B can interact with several annexin (ANX) proteins. The annexins are a family of proteins that bind multiple Ca2+ ions via Ca2+-binding sites that are not EF-hand motifs but are referred to as type II and type III Ca2+-binding sites [11]. The X-ray crystallographic analysis of ANXA2 established a single type II Ca2+-binding site in each of the second, third, and fourth domains and two type III Ca2+-binding sites in the first domain of the protein. The characteristic feature of all annexins is the presence of several copies of an annexin repeat, which is defined as an approximately 70-residue-long conserved structural element that is required for the Ca2+ and membrane-binding function of the annexins. The presence of such annexin repeats combined with the capability of a protein to interact with the negatively charged phospholipids in a Ca2+-dependent manner is the major criterion defining the family of annexins. There are 12 human annexins (ANXA1-ANXA11 and ANXA13). ANXA1, ANXA2, ANXA5, ANXA6, and ANXA11 interact with S100 proteins [12]. S100A1, S100A4, S100A6, S100A10, S100A11, S100A12, and S100B can interact with annexins. Interestingly, the S100 proteins and annexins can form multiple complexes. For example, ANXA2 is found in complexes with S100A4, S100A6, S100A10, and S100A11 [13]. However, the complex formation is much more favorable in the case of S100A10 (Kd of 13 nM) compared to S100A4 (Kd of 5 uM) [14]. Furthermore, although both S100A4 and S100A10 promote plasminogen activation [15,16], plasminogen activation by S100A4 but not S100A10 is Ca2+-dependent.
2. Mutualistic Symbiosis
Mutualistic symbiosis refers to the symbiotic relationship between individuals of different species in which both individuals benefit from the association. We propose that this biological relationship between species can be extended to the subunits within a protein complex. As will be discussed in detail, the formation of the ANXA2/S100A10 complex regulates the biological activity of each subunit and confers new biological functions that were not possessed by either subunit. Furthermore, complex formation is also critical for the survival of S100A10 because, in the absence of ANXA2, the protein is rapidly degraded [58].
2.1. Mutualistic Symbiosis I-Survival of S100A10
S100A10 is undetectable unless ANXA2 is also expressed [58,59,60,61,62]. Thus, S100A10 is an extrinsically disordered protein that is rapidly degraded [63]. However, the binding of S100A10 to DLC1 does not protect S100A10 from degradation [64]. However, ANXA2 competes with DLC1 for binding to S100A10, suggesting that the interaction of S100A10 with ANXA2 or DLC1 results in unique conformations of S100A10. Previous studies demonstrated that the depletion of ANXA2 from cells resulted in the rapid ubiquitylation and proteasomal degradation of S100A10. One of these studies utilized protein overexpression with site-directed S100A10 mutants to propose that the carboxyl-terminal residues, Lys-91 and/or Lys-93, function as the ubiquitylation sites [65]. However, these studies failed to directly confirm the presence of ubiquitylated S100A10 at these sites by mass spectrometric analysis. We have also observed that the forced expression of S100A10 and ubiquitin results in the ubiquitination of S100A10, and we identified, for the first time, that the ubiquitination of S100A10 in forced expression systems occurs on Lys-57 [66]. Our study also showed that the mutagenesis of Lys-57 prevents the ubiquitylation and degradation of overexpressed S100A10. In contrast, another group identified Lys-47 as a site of the succinylation of S100A10 and proposed that this site was also a site of ubiquitylation [67]. Interestingly, Lys-47, Lys-54, and Lys-57 were identified as sites of the ubiquitylation of S100A10 by the mass spectrometric analysis of tissue extracts [68], although these sites were not confirmed by site-directed mutagenesis. It was also not clear why only two of the five tissues examined in that study detected ubiquitinated S100A10. It is interesting to note that intrinsically disordered regions have been identified in S100A10 and that Lys-57 is located in the helix III, which is an intrinsically disordered region [63]. We analyzed the amino acid sequence of the S100A10 of twenty-one vertebrate species. Although Lys-57 is conserved in all vertebrate species, Lys-47 is not conserved in chickens, Xenopus or pufferfish.
2.2. Mutualistic Symbiosis II-Regulation of the Biological Functions of ANXA2 and S100A10
The mutualistic symbiotic role of S100A10 binding is apparent from studies of the regulation of F-actin bundling. Individually, S100A10 and ANXA2 are incapable of the significant bundling of F-actin; however, when ANXA2 bound S100A10, the complex facilitated the Ca2+-dependent formation of anisotropic F-actin bundles [43]. Because ANXA2 is an F-actin-binding protein, whereas S100A10 does not interact with F-actin, this type of mutualistic symbiosis involves modifying the function of one subunit by the other.
AIIt-dependent F-actin bundles participate in the spatial organization of the plasma membrane, providing active sites for secretory granule docking and exocytotic fusion. For example, it has been shown that when chromaffin cells are stimulated with nicotine, the F-actin-bundling activity of ANXA2 promotes the formation of the monosialotetrahexosylganglioside and GM1-enriched microdomains at the plasma membrane, increases the number of morphologically docked granules at the plasma membrane, and controls the number of individual exocytotic events [73]. Conversely, when an ANXA2 mutant with impaired F-actin filament–bundling activity [44] was expressed, the formation of plasma membrane lipid microdomains and the number of exocytotic events were decreased, and the fusion kinetics were slower [73]. Based on these studies, the authors concluded that AIIt-induced F-actin bundling is essential for generating active exocytotic sites.
Chasserot-Golaz’s group proposed a model for the involvement of AIIt in neurosecretion that incorporates both the F-actin bundling and the lipid sequestration properties of AIIt [74]. In unstimulated adrenergic cells, ANXA2 is cytosolic, S100A10 is bound to vesicle-associated membrane protein 2 (VAMP2) at the plasma membrane, and the membrane lipids PIP2 and GM1 are randomly distributed in the plasma membrane. However, upon stimulation with secretagogues, ANXA2 translocates to the plasma membrane in proximity to N-ethylmaleimide-sensitive factor attachment protein receptor (SNARE) complexes; this redistribution of ANXA2 is mediated by S100A10 [75]. The newly formed AIIt forms specific lipid microdomains. AIIt then bundles F-actin, which constrains the lateral mobility of the newly formed lipid domains, thereby increasing their stability. This compartmentalization of the plasma membrane serves to spatially and temporally organize F-actin bundles, which contribute to the docking machinery and thereby accelerate the vesicle fusion proteins required for secretory granule docking and fusion, likely playing a role in both the speed and accuracy of the exocytotic fusion process.
The biochemical properties of the monomeric ANXA2 and ANXA2 present in the AIIt complex are distinct. ANXA2 typically exists as a monomer predominantly localized to the cytosol and on early endosomes [76] or as a heterotetrameric complex—AIIt—found in the subplasmalemmal region [77]. Several laboratories have reported that the binding of S100A10 to ANXA2 induces a conformational change in ANXA2, which influences its interaction with several of its ligands. For example, S100A10 binding has a stimulatory effect on phospholipid binding by ANXA2, and the phospholipid requirement for 50% binding is 10-fold lower for the complex than ANXA2 [78]. We initially investigated the consequence of complex formation on the biological properties of the individual subunits by expressing and purifying recombinant ANXA2 and S100A10 and then combining the subunits and isolating AIIt. Previously, it was reported that only the ANXA2/S100A10 complex—and not monomeric ANXA2—could aggregate membrane vesicles at submicromolar Ca2+ concentrations [79,80]. We found that the binding of recombinant S100A10 to recombinant ANXA2 resulted in a decrease in the A0.5 (Ca2+) for chromaffin granule aggregation from 0.23 mM for recombinant ANXA2 to 1.0 μM for the recombinant complex. We also found that the binding of recombinant S100A10 to recombinant ANXA2 resulted in a decrease in the A0.5 (Ca2+) of phospholipid liposome aggregation from 0.83 μM for recombinant ANXA2 to 0.26 μM for the recombinant complex. In contrast, S100A10 did not interact with chromaffin granules or phospholipid vesicles.
The thioredoxin system reduces AIIt, enabling it to repeatedly exert its protein reductase activity. Because the mutagenesis of ANXA2 Cys-334-Ser and either S100A10 Cys-61-Ser or S100A10 Cys-82-Ser inactivated the plasmin reductase activity of the isolated subunits of AIIt, and because the sum of the plasmin reactivity (AIIt) is greater than the activity exhibited by each subunit, we concluded that each subunit acts in a mutualistic symbiotic relationship to enhance its plasmin reductase activity.
2.3. Mutualistic Symbiosis III—Acquisition of New Biological Activities by the Formation of the ANXA2/S100A10 Complex
The complex formed between ANXA2, S100A10, and AHNAK also interact with dysferlin [107] (Figure 4). This large multiprotein complex facilitates the wound repair of damaged epithelial, auditory, and muscle cells upon extracellular calcium influx [105,108,109,110]. Mechanistically, it has been suggested that this dysferlin membrane repair complex, when activated by calcium, promotes the fusion of exocytosis vesicles with the inner side of the plasma membrane, resulting in the resealing of membrane tears [111,112]. The ANXA2-mediated linking of membrane surfaces under non-oxidative intracellular conditions requires ANXA2-S100A10 complex formation because ANXA2 by itself cannot link biological membranes [80]. This suggests that the formation of the AIIt complex and its mutualistic symbiosis is required for membrane repair and that individually ANXA2 or S100A10 cannot participate in this process.
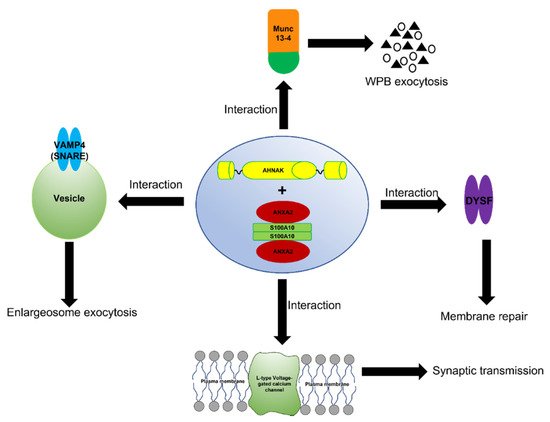
Figure 4. Schematic representation of the function of the AIIt-AHNAK complex. The formation of the ANXA2/S100A10/AHNAK (2:2:1) complex forms a key hub for the regulation of several important biological functions such as the exocytosis of enlargesomes, the regulation of the L-type voltage-gated calcium channel (VGCC), and possibly the MUNC-13-dependent regulation of the exocytosis of Weibel–Palade bodies (WPB).
The ANXA2/S100A10/AHNAK complex forms a complex with the L-type voltage-gated calcium channel (VGCC) [103], which plays a role in regulating Ca2+-dependent exocytosis. These channels are the primary regulators that control Ca2+ release and are pharmacological targets for treating cardiac ischemia and hypertension [113]. A decrease in the L-type calcium influx was observed in both the glutamatergic neurons and GABAergic interneurons of AHNAK-KO mice, suggesting that L-type calcium channels may act as effectors of the ANXA2/S100A10/AHNAK complex.
Recently, S100A10 was demonstrated to interact with the background potassium channel TASK-1 and regulate the membrane trafficking and the functionality of this channel (Girard et al., 2002). The tetrodotoxin-insensitive voltage-gated sodium channel, Nav1.8, has also been shown to bind S100A10 (Okuse et al., 2002). S100A10 promoted the translocation of Nav1.8 to the plasma membrane, thereby producing functional sodium channels. The role of ANXA2 has not been assessed for TASK-1 or Nav1.8. Therefore, it is unclear whether both ANXA2 and S100A10 are required for the activity of these ion channels.
Metabotropic glutamate receptors (mGluRs) are a sub-family of glutamate receptors. Previous studies have shown that antagonists acting on mGluR5 or mGluR2/3 exert antidepressant-like activities in mice [156]. Interestingly, an S100A10-binding motif has been identified in the cytoplasmic tail of mGluR5, and a C83Q S100A10 mutant which cannot interact with ANXA2, failed to interact with mGluR5. Therefore, the authors concluded that ANXA2 is required for the interaction of S100A10 with mGluR5 [157]. Accordingly, ANXA2 and S100A10 act in a mutualistic symbiotic relationship resulting in the regulation of mGluR5.
Mutations in CFTR, a cAMP/PKA, and the ATP-regulated Cl- channel result in the disease cystic fibrosis. AIIt forms a PKA/calcineurin-dependent complex with CFTR and tethers the complex to the plasma membrane [152]. Because the forced disruption of AIIt by a peptide to the S100A10 binding site disrupts the AIIt-CFTR complex and inhibits outwardly rectifying Cl− channels, the authors concluded that both subunits of AIIt were required for channel activity. Therefore, ANXA2 and S100A10 act in a mutualistic symbiotic relationship resulting in the regulation of CFTR.
Endosomes are subcellular organelles associated with the catabolism of exogenous and endogenous proteins and the down-regulation of surface receptors. They exist as three distinct cellular compartments: early endosomes, late endosomes, and recycling endosomes. S100A10 has been suggested to be dispensable for ANXA2 functions in transporting endocytosed tracers from early to late endosomes. The N-terminus of ANXA2 has a membrane-binding motif (amino acids 15 to 24) that regulates its Ca2+-independent association with endosomes [158]. The depletion of ANXA2 prevents newly-formed multivesicular endosomes from detaching from early endosomes and regulates their transport towards late endosomes [76]. However, AIIt plays a role in the subcellular distribution of early and recycling endosomes [62,159]. To account for these differences, it has been suggested that calcium-dependent membrane association at the plasma membrane or along the protein recycling pathway is regulated by S100A10 binding and AIIt formation [54]. For example, S100A10 appears necessary for ANXA2 binding to the plasma membrane and the cortical actin cytoskeleton. Utilizing HPV as model cargo and BSA as pathway-specific canonical cargo in uptake experiments, Taylor et al. [136] demonstrated that S100A10 and ANXA2 are required for endosomal vesicular trafficking. They also suggested that AIIt functions in the biogenesis of multivesicular endosomes. Therefore, ANXA2 and S100A10 act in a mutualistic symbiotic relationship to regulate certain aspects of the endosomal pathway.
In conclusion, ANXA2 and S100A10 collaborate in regulating multiple cellular functions, including specific ion channels. In some cases, S100A10 and ANXA2 provide binding sites for an ion channel, and in other cases, each subunit may play distinct but essential regulatory roles. In some instances, ANXA2 may tether the S100A10-ligand/channel complex to the plasma membrane. The ANXA2-S100A10-AHNAK complex also plays key roles in regulating L-type voltage-gated calcium channels and in the membrane repair process.
This entry is adapted from the peer-reviewed paper 10.3390/biom11121849
This entry is offline, you can click here to edit this entry!