Metabolic syndrome (MetS) is a non-communicable disease characterised by a cluster of metabolic irregularities. The prevalence of MetS in people living with Human Immunodeficiency Virus (HIV) and antiretroviral (ARV) usage is increasing rapidly. The findings suggest that mitochondrial dysfunction was the most common mechanism that induced metabolic complications. Furthermore, protease inhibitors (PIs) are more commonly implicated in MetS-related effects than other classes of ARVs.
- metabolic syndrome
- HIV
- ARVs
- mitochondrial dysfunction
- inflammation
1. Introduction
2. HIV and MetS
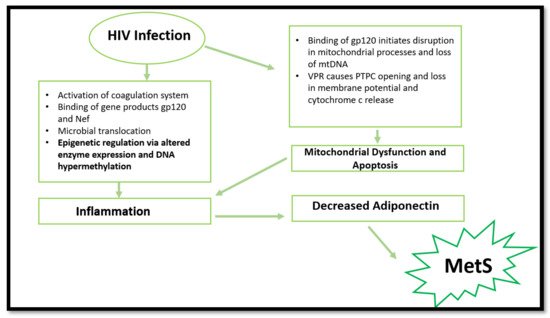
3. HIV, Mitochondrial Dysfunction, and Cell Apoptosis
4. HIV and Epigenetic Modifications
Aside from the common mechanisms, more recent research has suggested that HIV-1 infection can cause epigenetic changes when exposed to Mycobacterium tuberculosis. The latter results in altered monocyte function and dysregulation in pro-inflammatory cytokine production. The same study suggested that a decrease in global DNA methylation occurred in HIV-infected individuals. This was mainly attributed to the downregulation of DNA methyltransferases and the upregulation of methyl-CpG-binding proteins. Consequently, the reduction in global DNA methylation caused an increased activation status of monocytes. This result was accompanied by increased production of pro-inflammatory cytokines [16]. These findings are significant, considering the vulnerability to Mycobacterium tuberculosis in developing countries with high HIV prevalence, such as South Africa. The study provides plausible cause to initiate future in vivo research that will aid in understanding epigenetic changes associated with HIV and MetS.
More recent findings report DNA hypermethylation in HIV-infected patients. These epigenetic modifications result in cell dysfunction and decreased cytokine production, thus increasing CVD and T2DM risk [17]. More specifically, the HIV-1 infection causes epigenetic changes in the T-cell population, resulting in aberrant expression of pro-inflammatory cytokines and immune-related genes [18]. This is caused by the virus inducing DNA methylation changes in essential genes (IL-2, PD-1 and FOXP3) in T-cells which initiates cell dysfunction [19][20][21].
5. The Evolution of ARVs and Implications in MetS
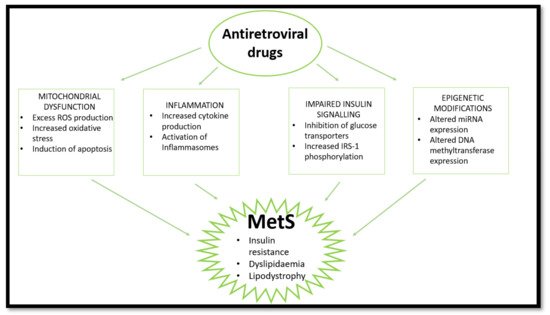
6. ARVs and Epigenetic Modifications—Emerging Evidence
This entry is adapted from the peer-reviewed paper 10.3390/ijms222112020
References
- Prasun, P. Mitochondrial dysfunction in metabolic syndrome. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2021, 1866, 165838.
- Saklayen, M.G. The global epidemic of the metabolic syndrome. Curr. Hypertens. Rep. 2018, 20, 1–8.
- Todowede, O.O.; Mianda, S.Z.; Sartorius, B. Prevalence of metabolic syndrome among HIV-positive and HIV-negative populations in sub-Saharan Africa—a systematic review and meta-analysis. Syst. Rev. 2019, 8, 4.
- UNAIDS. Global HIV & AIDS Statistics—2020 Fact Sheet. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 4 April 2021).
- Stats-SA. 2020 Mid-Year Population Estimates. Available online: http://www.statssa.gov.za/?p=13453 (accessed on 22 July 2021).
- Jaggers, J.R.; Prasad, V.K.; Dudgeon, W.D.; Blair, S.N.; Sui, X.; Burgess, S.; Hand, G.A. Associations between physical activity and sedentary time on components of metabolic syndrome among adults with HIV. AIDS Care 2014, 26, 1387–1392.
- Nguyen, K.; Peer, N.; Mills, E.J.; Kengne, A.P. A meta-analysis of the metabolic syndrome prevalence in the global HIV-infected population. PLoS ONE 2016, 11, e0150970.
- Syed, F.F.; Sani, M.U. Recent advances in HIV-associated cardiovascular diseases in Africa. Heart 2013, 99, 1146–1153.
- Masuku, S.K.S.; Tsoka-Gwegweni, J.; Sartorius, B. HIV and antiretroviral therapy-induced metabolic syndrome in people living with HIV and its implications for care: A critical review. J. Diabetol. 2019, 10, 41.
- Roda, R.H.; Hoke, A. Mitochondrial dysfunction in HIV-induced peripheral neuropathy. Int. Rev. Neurobiol. 2019, 145, 67–82.
- Grunfeld, C.; Pang, M.; Doerrler, W.; Shigenaga, J.; Jensen, P.; Feingold, K. Lipids, lipoproteins, triglyceride clearance, and cytokines in human immunodeficiency virus infection and the acquired immunodeficiency syndrome. J. Clin. Endocrinol. Metab. 1992, 74, 1045–1052.
- Haugaard, S.B.; Andersen, O.; Pedersen, S.B.; Dela, F.; Fenger, M.; Richelsen, B.; Madsbad, S.; Iversen, J. Tumor necrosis factor α is associated with insulin-mediated suppression of free fatty acids and net lipid oxidation in HIV-infected patients with lipodystrophy. Metabolism 2006, 55, 175–182.
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377.
- Karpe, F.; Dickmann, J.R.; Frayn, K.N. Fatty acids, obesity, and insulin resistance: Time for a reevaluation. Diabetes 2011, 60, 2441–2449.
- Maseko, T.S.; Masuku, S.K. The effect of HIV and ART on the development of hypertension and type 2 diabetes mellitus. J. Diabetes Metab. 2017, 8, 1–8.
- Espíndola, M.S.; Soares, L.S.; Galvão-Lima, L.J.; Zambuzi, F.A.; Cacemiro, M.C.; Brauer, V.S.; Marzocchi-Machado, C.M.; de Souza Gomes, M.; Amaral, L.R.; Martins-Filho, O.A. Epigenetic alterations are associated with monocyte immune dysfunctions in HIV-1 infection. Sci. Rep. 2018, 8, 1–14.
- Sundermann, E.E.; Hussain, M.A.; Moore, D.J.; Horvath, S.; Lin, D.T.; Kobor, M.S.; Levine, A. Inflammation-related genes are associated with epigenetic aging in HIV. J. Neurovirol. 2019, 25, 853–865.
- Bogoi, R.N.; de Pablo, A.; Valencia, E.; Martín-Carbonero, L.; Moreno, V.; Vilchez-Rueda, H.H.; Asensi, V.; Rodriguez, R.; Toledano, V.; Rodés, B. Expression profiling of chromatin-modifying enzymes and global DNA methylation in CD4+ T cells from patients with chronic HIV infection at different HIV control and progression states. Clin. Epigenetics 2018, 10, 1–10.
- Nakayama-Hosoya, K.; Ishida, T.; Youngblood, B.; Nakamura, H.; Hosoya, N.; Koga, M.; Koibuchi, T.; Iwamoto, A.; Kawana-Tachikawa, A. Epigenetic repression of interleukin 2 expression in senescent CD4+ T cells during chronic HIV type 1 infection. J. Infect. Dis. 2015, 211, 28–39.
- Youngblood, B.; Oestreich, K.J.; Ha, S.-J.; Duraiswamy, J.; Akondy, R.S.; West, E.E.; Wei, Z.; Lu, P.; Austin, J.W.; Riley, J.L. Chronic virus infection enforces demethylation of the locus that encodes PD-1 in antigen-specific CD8+ T cells. Immunity 2011, 35, 400–412.
- Milián, L.; Peris, J.E.; Gandía, P.; Andújar, I.; Pallardó, L.; Górriz, J.L.; Blas-García, A. Tenofovir-induced toxicity in renal proximal tubular epithelial cells: Involvement of mitochondria. Aids 2017, 31, 1679–1684.
- Paula, A.A.; Falcão, M.C.; Pacheco, A.G. Metabolic syndrome in HIV-infected individuals: Underlying mechanisms and epidemiological aspects. AIDS Res. Ther. 2013, 10, 32.
- Dau, B.; Holodniy, M. The relationship between HIV infection and cardiovascular disease. Curr. Cardiol. Rev. 2008, 4, 203–218.
- Muyanja, D.; Muzoora, C.; Muyingo, A.; Muyindike, W.; Siedner, M.J. High prevalence of metabolic syndrome and cardiovascular disease risk among people with HIV on stable ART in southwestern Uganda. AIDS Patient Care STDs 2016, 30, 4–10.
- Obirikorang, C.; Quaye, L.; Osei-Yeboah, J.; Odame, E.A.; Asare, I. Prevalence of metabolic syndrome among HIV-infected patients in Ghana: A cross-sectional study. Niger. Med. J. J. Niger. Med. Assoc. 2016, 57, 86.
- Hernández-Walias, F.; Ruiz-de-León, M.J.; Rosado-Sánchez, I.; Vázquez, E.; Leal, M.; Moreno, S.; Vidal, F.; Blanco, J.; Pacheco, Y.M.; Vallejo, A. New signatures of poor CD4 cell recovery after suppressive antiretroviral therapy in HIV-1-infected individuals: Involvement of miR-192, IL-6, sCD14 and miR-144. Sci. Rep. 2020, 10, 1–11.
- Bresciani, E.; Saletti, C.; Squillace, N.; Rizzi, L.; Molteni, L.; Meanti, R.; Omeljaniuk, R.J.; Biagini, G.; Gori, A.; Locatelli, V. miRNA-218 targets lipin-1 and glucose transporter type 4 genes in 3T3-L1 cells treated with lopinavir/ritonavir. Front. Pharmacol. 2019, 10, 461.
- Madeddu, G.; Ortu, S.; Garrucciu, G.; Maida, I.; Melis, M.; Muredda, A.A.; Mura, M.S.; Babudieri, S. DNMT1 modulation in chronic hepatitis B patients and hypothetic influence on mitochondrial DNA methylation status during long-term nucleo (t) side analogs therapy. J. Med. Virol. 2017, 89, 1208–1214.
- Yu, J.; Qiu, Y.; Yang, J.; Bian, S.; Chen, G.; Deng, M.; Kang, H.; Huang, L. DNMT1-PPARγ pathway in macrophages regulates chronic inflammation and atherosclerosis development in mice. Sci. Rep. 2016, 6, 1–11.
- Kim, A.Y.; Park, Y.J.; Pan, X.; Shin, K.C.; Kwak, S.-H.; Bassas, A.F.; Sallam, R.M.; Park, K.S.; Alfadda, A.A.; Xu, A. Obesity-induced DNA hypermethylation of the adiponectin gene mediates insulin resistance. Nat. Commun. 2015, 6, 1–11.