Drosophila melanogaster has proved to be a dynamic model organism that can produce high-quality data in a short time frame. One of the fly’s most prominent feature is the possibility to perform genetic alterations through the well-known Gal4/UAS expression system, thus making it possible to express target proteins in a specific cell type or tissue.
1. Introduction
Neurodegenerative diseases, such as Alzheimer’s disease (AD), are associated with proteotoxicity, which is caused by protein aggregation and results in extensive neuronal damage in the brain. Neurodegeneration and the disturbance of essential functions in the cell manifest in cognitive impairments and premature death [1,2]. A main proteotoxic contributor in AD is the amyloid-β (Aβ) peptide, where the propensity to aggregate differs between different variants [3]. Aβ misfolds and aggregates into pre-fibrillar assemblies that are highly associated with toxicity. They then progressively merge into mature fibrils [1,4]. There is an urgent need to find disease-modifying treatments and, therefore, reliable and powerful methods to pin down fundamental components underlying the mechanisms responsible for proteotoxicity are needed. Drosophila offers a number of advantages as a model system for studying diseases where: (i) a variety of phenotypic markers are available for identifying detrimental effects due to proteotoxicity, (ii) the lifespan of Drosophila makes it possible to investigate age-related diseases on a reasonable time scale (days to weeks, as opposed to months and years, in mouse model systems), (iii) the system is amenable to large drug screens since the flies proliferate well and are relatively inexpensive and easy to work with and (vi) there are extensive tools that allow disease-related genes and molecular pathways to be genetically and pharmacologically manipulated in order to find out both the function of their orthologs in vivo, and how these genes are involved in the pathogenesis of different diseases, which can generate in vivo data that are translatable to mammalian system [6,7].
2. Drosophila as Model Organism for Drug Screen against Aβ Proteotoxicity
There are two main approaches to delivering the drug when using Drosophila to screen for compounds against proteotoxicity. The drug molecule can be mixed in the food and administrated to the fly expressing the proteotoxic protein, or, if the drug is a protein, it can be co-expressed with the proteotoxic protein in the fly.
2.1. Blocking Aβ Aggregation
In one of the first
Drosophila models of AD, the amyloid-binding dye Congo Red was tested for protection against the proteotoxicity of the Aβ peptide [
15]. Feeding the flies food mixed with Congo Red extended the life span of both Aβ1-42 and Arctic (Glu22Gly) Aβ1-42-expressing flies. Histology experiments revealed fewer protein aggregates in Aβ1-42-expressing flies treated with Congo Red compared to untreated flies. This study shows that the proteotoxic effect of the Aβ peptide can be hindered in vivo by a drug that blocks the aggregation process of the peptide. Indeed, considering the connection between the formation of toxic Aβ species and AD, finding a drug that inhibits the Aβ aggregation process should be an effective therapeutic strategy. Using this approach, a synthetic molecule designated D737 (C25H20N2O) was discovered when a library of compounds was screened for anti-Aβ aggregation properties [
50]. The compound D737 increased the life span and improved the climbing performance of both Aβ1-42 and Arctic Aβ1-42-expressing flies [
51]. In a follow up study, two analogs of D737 with anti-Aβ aggregation properties (D744 and D830) were identified. These analogues were able to rescue Aβ proteotoxicity more efficiently than D737, thus strengthening the evidence of a correlation between the anti-Aβ aggregation properties of a drug and its ability to block Aβ toxicity in vivo. Another study found that feeding AD flies with acetylcholinesterase inhibitors improved the longevity and mobility of Arctic (Glu22Gly) Aβ1-42 flies and that the number of aggregates in the fly brain was reduced [
52]. This rescue effect was particularly evident for the newly synthesized acetylcholinesterase inhibitor XJP-1 and was attributed to the ability of XJP-1 to inhibit the acetylcholinesterase-induced aggregation of the Aβ peptide.
2.2. Enhancing Aβ Aggregation
In contrast, when the effect of curcumin on Aβ proteotoxicity was investigated in
Drosophila, it was found that the toxicity of the Aβ peptide can be suppressed by enhancing the fibrillation process [
27]. In this study, flies overexpressing Aβ1-42 or the Arctic (Glu22Gly) Aβ1-42 variant were fed with a substrate mixture containing curcumin. Although it was found that curcumin by itself is somewhat toxic to the flies, survival and locomotor analyses showed a rescue effect for the Aβ expressing flies treated with curcumin. Histochemistry analyses revealed that the presence of curcumin accelerated the Aβ fibrillation process in the fly brain, thereby reducing the pool of toxic prefibrillar species. In non-transgenic
Drosophila, curcumin has shown to both down-regulate the gene expression of acetylcholinesterase, thereby increasing neuronal signaling, and to improve the antioxidant status [
53]. These factors might contribute to the anti-toxic effect of curcumin in AD flies.
2.3. Increasing Protein Clearance
A compound that can increase the ability of the cells to degrade the Aβ peptide would be an interesting drug candidate. Indeed, this year, the FDA approved aducanumab, the first monoclonal antibody treatment for AD (Biogen). This approach has also been investigated in
Drosophila, where an engineered Aβ binding affibody protein was co-expressed with the Aβ peptide in the fly brain [
54]. The affibody molecule consists of a three-helix Z domain and can be selected for different binding properties using phage display. The presence of two copies of the affibody protein, connected head-to-tail, resulted in an impressive increase in the lifespan for both Aβ1-42 and Arctic (Glu22Gly) Aβ1-42-expressing flies. In addition, the abnormal rough eye phenotype of the Arctic Aβ1-42-expressing flies was suppressed. Biochemical analyses showed that the Aβ levels and deposits of Aβ aggregates in the fly brain decreased sharply, indicating that the anti-Aβ proteotoxic effect of the affibody protein is due to its ability to promote the clearance of the peptide in fly tissue. In a recent study, Aβ1-42-expressing flies were fed with extract from red adzuki beans [
55]. Data from this study showed a rescue effect for the Aβ flies, which manifested in an increased longevity and locomotor activity and an in memory improvement of the adzuki-bean-treated flies. In addition, the Aβ level of the treated Aβ flies decreased compared to the untreated flies, indicating that the intake of red adzuki beans improves the degradation process of the Aβ peptide in the fly brain, which protects the neurons against Aβ proteotoxicity. A similar result was found for the protein puromycin-sensitive aminopeptidase (SPA) that was tested for anti-toxic effects in AD flies [
56]. The co-expression of this enzyme in Arctic Aβ1-42 flies resulted in an increased life span and activity and the Aβ-induced rough eye phenotype was rescued. Additionally, the Aβ levels and deposits of Aβ in the fly brain were greatly reduced in the presence of SPA, suggesting that the rescue effect is due to the enzyme’s ability to enhance clearance of the peptide.
2.4. Proteins and Peptides as Drug Candidates
The advantage of testing a protein as an anti-proteotoxic drug in fly models is that the drug–protein can be co-expressed with the proteotoxic protein, ensuring that the two molecules will be present in the fly tissue simultaneously. This way, the lysozyme protein was tested for its anti-Aβ proteotoxic effect in AD flies [
57,
58]. Co-expressing lysozyme with Aβ1-42 extended the life span and improved the activity of the flies. In addition, the rough eye phenotype in Aβ1-42-expressing flies was suppressed. Lysozyme was found to interact with the Aβ1-42 peptide in vivo and to reduce the Aβ levels in the fly brain. These data suggest that the anti-toxic effect of lysozyme is due to its ability to disrupt the Aβ aggregation process, resulting in non-toxic species, and facilitating the Aβ degradation process.
Interestingly, shorter Aβ peptides can counteract Aβ proteotoxicity [
19]. The co-expression of Aβ1-36 to Aβ1-39 peptides with Aβ1-42 did partially rescue the locomotor dysfunction and rough eye phenotype of Aβ1-42 flies. The distribution of Aβ assemblies in the mushroom body neurons of the flies was not affected by the presence of the shorter peptides and no apparent correlation was found between their rescue effects and the Aβ1-42 levels in the flies. A proposed protection mechanism for these shorter peptides is that they interfere with the Aβ1-42 aggregation process in various ways, with the common result that the level of toxic Aβ species in the fly is reduced.
Overexpressing the chaperon domain proSP-C BRICHOS was found to protect AD flies from Aβ proteotoxicity [
59,
60]. The rescue effects were manifested in the extended lifespan and increased locomotor activity when proSP-C BRICHOS was co-expressed with Aβ1-42 in the fly neurons. In addition, the deposition of Aβ aggregates in the fly brain was delayed and the ratio between soluble and insoluble Aβ was increased. Later, a study was published where the anti-Aβ proteotoxic effects of the proSP-C BRICHOS and Bri2 BRICHOS domains were investigated in parallel. The study showed that the Bri2 BRICHOS domain can prevent Aβ1-42 toxicity in the flies in a similar fashion as the proSP-C BRICHOS domain, albeit more efficiently. Additionally, a rescue effect of the eye phenotype was also confirmed. In vitro analyses revealed that the BRICHOS domains inhibit the aggregation process of Aβ but in different ways. Whereas proSP-C BRICHOS specifically affects the secondary nucleation event [
61], Bri2 BRICHOS inhibits the aggregation in a more comprehensive way, affecting both the secondary nucleation and fibril-end elongation. Both BRICHOS domains interfere with the Aβ aggregation in such a way that the formation of toxic Aβ species is reduced, which slows down the disease progression in the flies.
2.5. Targeting Inflammatory Processes
It is well known that inflammation is a prominent feature in AD [
62]. Thus, one therapeutic strategy is to find natural or synthetic drugs with anti-inflammatory properties. With this in mind, Aβ1-42-expressing flies were treated with an extract from the plant
Arabidopsis thaliana known to contain polyphenols, which is a group of natural compounds that possess anti-inflammatory properties [
63]. The extract was found to increase the activity of the flies when assessed by a climbing assay. Among the polyphenol compounds that were detected in the extract from
Arabidopsis thaliana, two derivates of kaempferol and quercetin were identified, as well as luteolin. These substances have been tested separately for their anti-Aβ proteotoxic effect in AD flies. Quercetin was found to extend the lifespan and increase the activity of Arctic (Glu22Gly) Aβ1-42-expressing flies [
64]. In this study, a detailed transcriptomic analysis revealed the disturbance of cell signaling pathways in the AD flies. Specifically, the expression of proteins involved in the FoxO cell cycle signaling pathway and in DNA replication was found to be dysregulated in the AD flies, which most likely contributes to toxicity. These pathways were largely restored by the presence of quercetin in the fly brain, which indicates that the anti-Aβ proteotoxicity mechanism can be attributed to the compound’s ability to re-establish cell signaling pathways and DNA replication. Feeding Aβ1-42-expressing flies with kaempferol increased the climbing performance and protected the AD flies from memory loss and oxidative stress. In addition, the compound reduced the acetylcholinesterase activity, which increases neuronal signaling [
65]. Luteolin was found to rescue Aβ1-42 toxicity in longevity and climbing assays, and the formation of Aβ aggregates in the AD fly brain was reduced [
66]. Moreover, the acetylcholinesterase activity and oxidative stress were suppressed in the luteolin-treated AD flies. In summary, in addition to anti-inflammatory characteristics, the protection of
Arabidopsis thaliana extract against Aβ toxicity is likely due to a combination of anti-Aβ proteotoxic effects exerted by various polyphenols, which includes a decrease in the formation of Aβ aggregates, a decrease in acetylcholinesterase activity, the restoration of cell signaling pathways for the cell cycle and DNA replication and protection against oxidative stress.
Treating Aβ1-42-expressing flies with extracts from
Gardenia jasminoides did also rescue Aβ toxicity without any detectable interfering with the Aβ aggregation process [
67]. The anti-proteotoxic effect of the extract was manifested by preventing memory loss of the AD flies. Quantifying the level of soluble and insoluble Aβ levels in the flies did not show any differences between treated and non-treated Aβ flies, leading to the conclusion that the rescue effect was not due to any changes in the Aβ aggregation process. Instead, the study showed that the rescue effect of the extract was due to its capacity to downregulate the expression of inflammatory genes that were found to be upregulated, causing toxicity in the Aβ flies.
2.6. Preventing Oxidative Stress
Reducing oxidative stress could be a very important target for a therapeutic strategy against AD, since the disease is thought to be accompanied by an excessive production of ROS, leading to cell death [
24]. Interestingly, feeding Aβ1-42-expressing flies with nordihydroguaiaretic acid (NDGA), which possesses both antioxidant and free radical scavenging properties, extended the life span and increased the climbing ability of the Aβ flies [
68]. A delay in the memory loss was also detected for the NDGA-treated Aβ flies, and oxidative stress and the acetylcholinesterase activity were reduced. The deposition of Aβ in the fly brain was not affected by the presence of NDGA, which indicates that the anti-Aβ proteotoxic effect of NDGA does not involve the disruption of the Aβ aggregation process but is rather exerted by the ability of NDGA to both increase neuronal signaling and reduce the formation of ROS in the AD flies.
2.7. Preventing Mitochondrial Dysfunction
Mitochondrial dysfunction is associated with AD [
69] and could thus be a relevant target when developing therapeutic strategies [
37]. This area has been explored in a study where AD flies that overexpressed a tandem variant of the Aβ1-42 peptide were treated with a compound named GMP-1 that is able to counteract mitochondrial dysfunction [
70]. Expressing a tandem repeat of the Aβ1-42 peptide increases the ability of the peptide to form oligomeric aggregates and boosts the toxic effect in the flies [
45]. Using this AD model, the GMP-1 compound was tested for anti-Aβ proteotoxic effects in vivo. A neuroprotective effect was detected where the longevity and climbing behavior were improved for the GMP-1 treated flies, which was attributed to the ability of GMP-1 to restore the mitochondrial function in the AD flies.
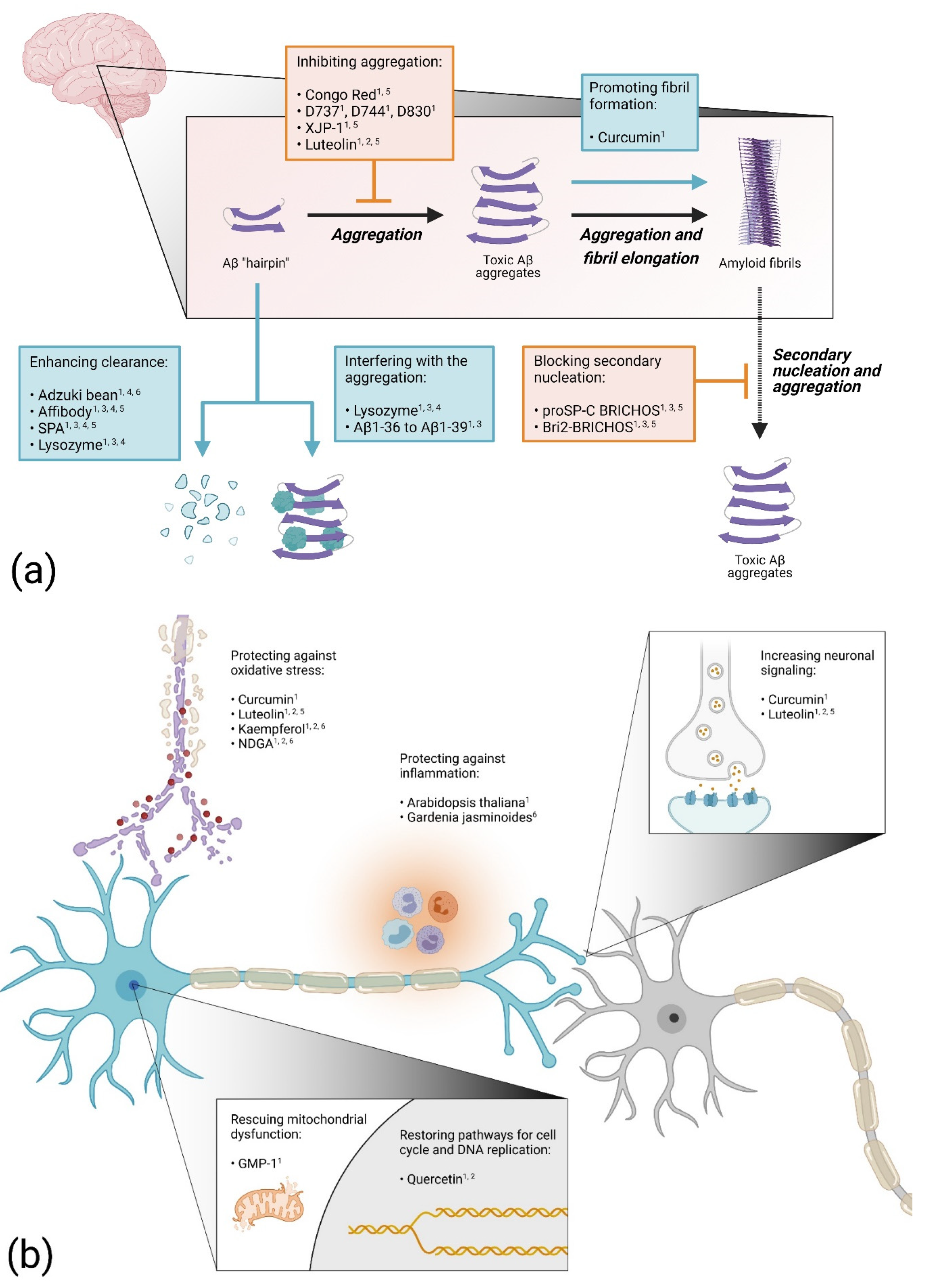
Figure 3 shows an overview of the different protective mechanisms of drugs with an anti-Aβ proteotoxic effect in Drosophila. These drug tests in Drosophila reveal that there might be several approaches to finding a treatment for AD. Most likely, a mixture of drugs with different modes of action would be necessary to block the disease progress and to ultimately cure the disease.
Figure 3. Overview of suggested anti-Aβ proteotoxicity mechanisms for different drugs examined in Drosophila: (a) drugs acting on the aggregation process; (b) drugs acting on cellular functions. Elevated numbers indicate confirmed protected effect in Alzheimer’s disease (AD) flies: 1 Improved viability; 2 Prevented adverse cellular impact; 3 Blocked cell death; 4 Reduced protein levels; 5 Reduced protein deposits; 6 Improved cognition. The figure was generated using BioRender.
3. Conclusions
Over the past two decades, Drosophila has been extensively used to study the disease mechanism behind protein aggregation and neurodegeneration for several protein misfolding diseases. In this review, we have focused on the use of AD fly models to investigate the proteotoxic effects of different Aβ isoforms, as well as to search for compounds that can counteract this toxicity. Although there are differences in the proteome between a fly brain and a human brain that might limit the possibility of directly applying results from fly experiments to humans, the fly has proven to be a powerful tool to unravel Aβ-related proteotoxicity and to find potential drug candidates. Thus, data from various fly studies of Aβ proteotoxicity are likely to make a significant contribution to ultimately finding a therapeutic strategy to cure AD.
This entry is adapted from the peer-reviewed paper 10.3390/ijms221910448