Tetraspanin 8 (TSPAN8) is a member of the tetraspanin superfamily that forms TSPAN8-mediated protein complexes by interacting with themselves and other various cellular signaling molecules. These protein complexes help build tetraspanin-enriched microdomains (TEMs) that efficiently mediate intracellular signal transduction. In physiological conditions, TSPAN8 plays a vital role in the regulation of biological functions, including leukocyte trafficking, angiogenesis and wound repair. Recently, reports have increasingly shown the functional role and clinical relevance of TSPAN8 overexpression in the progression and metastasis of several cancers.
- tetraspanin 8
- prognostic marker
- therapeutic target
- cancer
1. Introduction
2. The Structure and Physiological Role of TSPAN8 in Cells
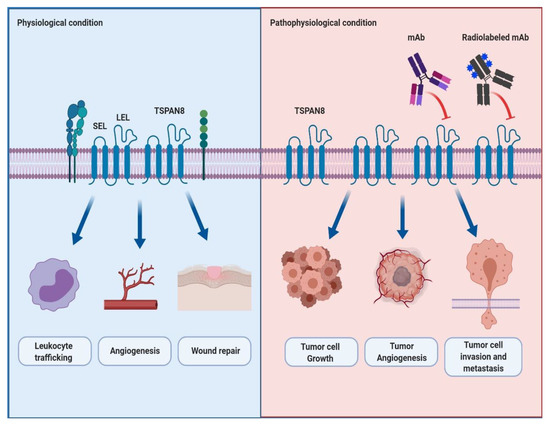
3. The Role of TSPAN8 in Cancer Progression and Metastasis
3.1. Pancreatic Cancer
3.2. Colon Cancers
3.3. Gastric Cancers
3.4. Liver Cancers
3.5. Lung Cancers
3.6. Breast Cancers
3.7. Ovarian Cancers
3.8. Gliomas
3.9. Melanomas
3.10. Esophageal Cancers
3.11. Nasopharyngeal Cancers
3.12. Cancer Stem Cells
This entry is adapted from the peer-reviewed paper 10.3390/biom10030388
References
- Singh, S.; Kumar, N.K.; Dwiwedi, P.; Charan, J.; Kaur, R.; Sidhu, P.; Chugh, V.K. Monoclonal Antibodies: A Review. Curr. Clin. Pharmacol. 2018, 13, 85–99.
- Kaplon, H.; Muralidharan, M.; Schneider, Z.; Reichert, J.M. Antibodies to watch in 2020. MAbs 2020, 12, 1703531.
- Monoclonal antibody approved for metastatic breast cancer. Oncology 1998, 12, 1727.
- Bardelli, A.; Siena, S. Molecular mechanisms of resistance to cetuximab and panitumumab in colorectal cancer. J. Clin. Oncol. 2010, 28, 1254–1261.
- Friedl, P.; Wolf, K. Tumour-cell invasion and migration: Diversity and escape mechanisms. Nat. Rev. Cancer 2003, 3, 362–374.
- Zoller, M. Tetraspanins: Push and pull in suppressing and promoting metastasis. Nat. Rev. Cancer 2009, 9, 40–55.
- Bonnet, M.; Maisonial-Besset, A.; Zhu, Y.; Witkowski, T.; Roche, G.; Boucheix, C.; Greco, C.; Degoul, F. Targeting the Tetraspanins with Monoclonal Antibodies in Oncology: Focus on Tspan8/Co-029. Cancers 2019, 11, 179.
- Szala, S.; Kasai, Y.; Steplewski, Z.; Rodeck, U.; Koprowski, H.; Linnenbach, A.J. Molecular cloning of cDNA for the human tumor-associated antigen CO-029 and identification of related transmembrane antigens. Proc. Natl. Acad. Sci. USA 1990, 87, 6833–6837.
- Claas, C.; Seiter, S.; Claas, A.; Savelyeva, L.; Schwab, M.; Zoller, M. Association between the rat homologue of CO-029, a metastasis-associated tetraspanin molecule and consumption coagulopathy. J. Cell Biol. 1998, 141, 267–280.
- Champy, M.F.; Le Voci, L.; Selloum, M.; Peterson, L.B.; Cumiskey, A.M.; Blom, D. Reduced body weight in male Tspan8-deficient mice. Int. J. Obes. 2011, 35, 605–617.
- Zhao, K.; Erb, U.; Hackert, T.; Zoller, M.; Yue, S. Distorted leukocyte migration, angiogenesis, wound repair and metastasis in Tspan8 and Tspan8/CD151 double knockout mice indicate complementary activities of Tspan8 and CD51. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865, 379–391.
- Gesierich, S.; Paret, C.; Hildebrand, D.; Weitz, J.; Zgraggen, K.; Schmitz-Winnenthal, F.H.; Horejsi, V.; Yoshie, O.; Herlyn, D.; Ashman, L.K.; et al. Colocalization of the tetraspanins, CO-029 and CD151, with integrins in human pancreatic adenocarcinoma: Impact on cell motility. Clin. Cancer Res. 2005, 11, 2840–2852.
- Yue, S.; Mu, W.; Zoller, M. Tspan8 and CD151 promote metastasis by distinct mechanisms. Eur. J. Cancer 2013, 49, 2934–2948.
- Claas, C.; Wahl, J.; Orlicky, D.J.; Karaduman, H.; Schnolzer, M.; Kempf, T.; Zoller, M. The tetraspanin D6.1A and its molecular partners on rat carcinoma cells. Biochem. J. 2005, 389, 99–110.
- Gesierich, S.; Berezovskiy, I.; Ryschich, E.; Zoller, M. Systemic induction of the angiogenesis switch by the tetraspanin D6.1A/CO-029. Cancer Res. 2006, 66, 7083–7094.
- Greco, C.; Bralet, M.P.; Ailane, N.; Dubart-Kupperschmitt, A.; Rubinstein, E.; Le Naour, F.; Boucheix, C. E-cadherin/p120-catenin and tetraspanin Co-029 cooperate for cell motility control in human colon carcinoma. Cancer Res. 2010, 70, 7674–7683.
- Guo, Q.; Xia, B.; Zhang, F.; Richardson, M.M.; Li, M.; Zhang, J.S.; Chen, F.; Zhang, X.A. Tetraspanin CO-029 inhibits colorectal cancer cell movement by deregulating cell-matrix and cell-cell adhesions. PLoS ONE 2012, 7, e38464.
- Anami, K.; Oue, N.; Noguchi, T.; Sakamoto, N.; Sentani, K.; Hayashi, T.; Naito, Y.; Oo, H.Z.; Yasui, W. TSPAN8, identified by Escherichia coli ampicillin secretion trap, is associated with cell growth and invasion in gastric cancer. Gastric Cancer 2016, 19, 370–380.
- Zhu, H.; Wu, Y.; Zheng, W.; Lu, S. CO-029 is overexpressed in gastric cancer and mediates the effects of EGF on gastric cancer cell proliferation and invasion. Int. J. Mol. Med. 2015, 35, 798–802.
- Wei, L.; Li, Y.; Suo, Z. TSPAN8 promotes gastric cancer growth and metastasis via ERK MAPK pathway. Int. J. Clin. Exp. Med. 2015, 8, 8599–8607.
- Li, L.; Yang, D.; Cui, D.; Li, Y.; Nie, Z.; Wang, J.; Liang, L. Quantitative proteomics analysis of the role of tetraspanin-8 in the drug resistance of gastric cancer. Int. J. Oncol. 2018, 52, 473–484.
- Kanetaka, K.; Sakamoto, M.; Yamamoto, Y.; Yamasaki, S.; Lanza, F.; Kanematsu, T.; Hirohashi, S. Overexpression of tetraspanin CO-029 in hepatocellular carcinoma. J. Hepatol. 2001, 35, 637–642.
- Fang, T.; Lin, J.; Wang, Y.; Chen, G.; Huang, J.; Chen, J.; Zhao, Y.; Sun, R.; Liang, C.; Liu, B. Tetraspanin-8 promotes hepatocellular carcinoma metastasis by increasing ADAM12m expression. Oncotarget 2016, 7, 40630–40643.
- Akiel, M.A.; Santhekadur, P.K.; Mendoza, R.G.; Siddiq, A.; Fisher, P.B.; Sarkar, D. Tetraspanin 8 mediates AEG-1-induced invasion and metastasis in hepatocellular carcinoma cells. FEBS Lett. 2016, 590, 2700–2708.
- Dong, Z.; Zhao, L.; Lu, S.; Xiong, J.; Geng, Z. Overexpression of TSPAN8 Promotes Tumor Cell Viability and Proliferation in Nonsmall Cell Lung Cancer. Cancer Biother. Radiopharm. 2016, 31, 353–359.
- Voglstaetter, M.; Thomsen, A.R.; Nouvel, J.; Koch, A.; Jank, P.; Navarro, E.G.; Gainey-Schleicher, T.; Khanduri, R.; Gross, A.; Rossner, F.; et al. Tspan8 is expressed in breast cancer and regulates E-cadherin/catenin signalling and metastasis accompanied by increased circulating extracellular vesicles. J. Pathol. 2019, 248, 421–437.
- Park, C.S.; Kim, T.K.; Kim, H.G.; Kim, Y.J.; Jeoung, M.H.; Lee, W.R.; Go, N.K.; Heo, K.; Lee, S. Therapeutic targeting of tetraspanin8 in epithelial ovarian cancer invasion and metastasis. Oncogene 2016, 35, 4540–4548.
- Pan, S.J.; Wu, Y.B.; Cai, S.; Pan, Y.X.; Liu, W.; Bian, L.G.; Sun, B.; Sun, Q.F. Over-expression of tetraspanin 8 in malignant glioma regulates tumor cell progression. Biochem. Biophys. Res. Commun. 2015, 458, 476–482.
- Pan, S.J.; Zhan, S.K.; Pan, Y.X.; Liu, W.; Bian, L.G.; Sun, B.; Sun, Q.F. Tetraspanin 8-rictor-integrin alpha3 complex is required for glioma cell migration. Int. J. Mol. Sci. 2015, 16, 5363–5374.
- Jiang, H.; Liu, W.; Zhan, S.K.; Pan, Y.X.; Bian, L.G.; Sun, B.; Sun, Q.F.; Pan, S.J. GSK621 Targets Glioma Cells via Activating AMP-Activated Protein Kinase Signalings. PLoS ONE 2016, 11, e0161017.
- Berthier-Vergnes, O.; El Kharbili, M.; de la Fouchardiere, A.; Pointecouteau, T.; Verrando, P.; Wierinckx, A.; Lachuer, J.; Le Naour, F.; Lamartine, J. Gene expression profiles of human melanoma cells with different invasive potential reveal TSPAN8 as a novel mediator of invasion. Br. J. Cancer 2011, 104, 155–165.
- Agaesse, G.; Barbollat-Boutrand, L.; Sulpice, E.; Bhajun, R.; El Kharbili, M.; Berthier-Vergnes, O.; Degoul, F.; de la Fouchardiere, A.; Berger, E.; Voeltzel, T.; et al. A large-scale RNAi screen identifies LCMR1 as a critical regulator of Tspan8-mediated melanoma invasion. Oncogene 2017, 36, 446–457.
- El Kharbili, M.; Robert, C.; Witkowski, T.; Danty-Berger, E.; Barbollat-Boutrand, L.; Masse, I.; Gadot, N.; de la Fouchardiere, A.; McDonald, P.C.; Dedhar, S.; et al. Tetraspanin 8 is a novel regulator of ILK-driven beta1 integrin adhesion and signaling in invasive melanoma cells. Oncotarget 2017, 8, 17140–17155.
- Agaesse, G.; Barbollat-Boutrand, L.; El Kharbili, M.; Berthier-Vergnes, O.; Masse, I. p53 targets TSPAN8 to prevent invasion in melanoma cells. Oncogenesis 2017, 6, e309.
- El Kharbili, M.; Agaesse, G.; Barbollat-Boutrand, L.; Pommier, R.M.; de la Fouchardiere, A.; Larue, L.; Caramel, J.; Puisieux, A.; Berthier-Vergnes, O.; Masse, I. Tspan8-beta-catenin positive feedback loop promotes melanoma invasion. Oncogene 2019, 38, 3781–3793.
- Zhou, Z.; Ran, Y.L.; Hu, H.; Pan, J.; Li, Z.F.; Chen, L.Z.; Sun, L.C.; Peng, L.; Zhao, X.L.; Yu, L.; et al. TM4SF3 promotes esophageal carcinoma metastasis via upregulating ADAM12m expression. Clin. Exp. Metastasis 2008, 25, 537–548.
- Lin, X.; Bi, Z.; Hu, Q.; Li, Q.; Liu, J.; Luo, M.L.; Xiang, Y.; Yao, H. TSPAN8 serves as a prognostic marker involving Akt/MAPK pathway in nasopharyngeal carcinoma. Ann. Transl. Med. 2019, 7, 470.
- Zhu, R.; Gires, O.; Zhu, L.; Liu, J.; Li, J.; Yang, H.; Ju, G.; Huang, J.; Ge, W.; Chen, Y.; et al. TSPAN8 promotes cancer cell stemness via activation of sonic Hedgehog signaling. Nat. Commun. 2019, 10, 2863.