-
- Bifidobacteria
1. History and Taxonomy
The Actinobacteria phylum represents one of the most numerous and heterogeneous groups of microorganisms present in nature [1,2]. These Gram-positive bacteria are characterized by a high GC genome content ranging from 51% to more than 70%, and exhibit different morphologies, including unicellular rods or Y-shaped rods, and complex multicellular consortia [1,2]. Furthermore, these bacteria are able to produce bioactive natural compounds and these features are reflected in their ability to adapt to several quite distinct ecosystems such as various terrestrial environments, as well as the bodies of mammals and birds [1,2]. In fact, this phylum includes pathogens (e.g., Mycobacterium spp., Nocardia spp., Tropheryma spp., Corynebacterium spp., and Propionibacterium spp.), soil inhabitants such as Streptomyces spp., plant commensals (e.g., Leifsonia spp.), nitrogen-fixing symbionts (Frankia), and human gut inhabitants (Bifidobacterium spp.) [1,2].
The genus Bifidobacterium belongs to the Bifidobacteriaceae family, Bifidobacteriales order, and these bacteria were isolated, for the first time, from feces of a breast-fed infant by Tissier in 1899 [3]. They represent nonmotile, anaerobic, nonsporulating, saccharolytic bacteria with a bifid or multiple-branching rod morphology. Currently, the genus Bifidobacterium comprises 94 taxa, representing 82 species and 12 subspecies [4-12] (Table 1). In recent years, the phylogeny of the Bifidobacterium genus has been explored using different methods based on the sequencing of the 16S rRNA gene, by means of a multilocus approach, or the sequencing of several housekeeping genes (i.e., clpC, dnaJ, rpoC, xpf, dnaB, and purF) [13,14]. A comparative genomics analysis based on all 88 sequenced bifidobacterial type strains revealed the presence of 191 Bifidobacterium-specific clusters of orthologous genes (COGs) shared by these genomes, called the bifidobacterial core-genome [15]. Notably, the phylogenetic tree constructed by amino acid concatenation of these 191 bifidobacterial core-genome genes revealed the existence of 10 different phylogenetic groups, encompassing Bifidobacterium adolescentis, Bifidobacterium boum, Bifidobacterium pullorum, Bifidobacterium asteroides, Bifidobacterium longum, Bifidobacterium psychraerophilum, Bifidobacterium bifidum, Bifidobacterium pseudolongum, Bifidobacterium bombi, and Bifidobacterium tissieri groups [15]. These groups partially correlate with the ecological niches from which the representative species were isolated. For example, members of the B. tissieri group are common inhabitants of the microbiota of tamarin and those of the B. pullorum group are characteristic of birds. According to this, members of the B. adolescentis group (Bifidobacterium catenulatum, Bifidobacterium pseudocatenulatum, and B. adolescentis strains), the B. longum group (Bifidobacterium breve and B. longum strains), the B. pseudolongum group (especially Bifidobacterium animalis subsp. lactis strains), and the B. bifidum group (B. bifidum strains) are typical colonizers of the human intestinal tract or are commercially exploited as probiotic strains (Figure 1).
Table 1. Bifidobacterium (sub)species recognized as reference strains (type strains).
|
Bifidobacterium Strains |
Isolation |
References |
|
B. actinocoloniiforme DSM 22766 |
Bumblebee digestive tract |
[16] |
|
B. adolescentis ATCC 15703 |
Intestine of human adult |
[17] |
|
B. aemilianum XV10 |
Carpenter bee digestive tract |
[5] |
|
B. aerophilum DSM 100689 |
Feces of cotton-top tamarin |
[18] |
|
B. aesuclapii DSM 26737 |
Feces of baby common marmoset |
[19] |
|
B. angulatum LMG 11039 |
Feces of human |
[20] |
|
B. animalis subsp. animalis LMG 10508 |
Feces of rat |
[21] |
|
B. animalis subsp. lactis DSM 10140 |
Fermented milk |
[22] |
|
B. anseris LMG 30189 |
Feces of domestic goose |
[7] |
|
B. apri DSM 100238 |
Digestive tract of wild pig |
[23] |
|
B. aquikefiri LMG 28769 |
Water kefir |
[24] |
|
B. asteroides LMG 10735 |
Hindgut of honeybee |
[25] |
|
B. avesanii DSM 100685 |
Feces of cotton-top tamarin |
[18] |
|
B. biavatii DSM 23969 |
Feces of tamarin |
[26] |
|
B. bifidum LMG 11041 |
Feces of breast-fed infant |
[3] |
|
B. bohemicum DSM22767 |
Bumblebee digestive tract |
[16] |
|
B. bombi DSM 19703 |
Bumblebee digestive tract |
[27] |
|
B. boum LMG 10736 |
Rumen of bovine |
[28] |
|
B. breve LMG 13208 |
Infant stool |
[17] |
|
B. callimiconis LMG 30938 |
Feces of Goeldi’s marmoset |
[6] |
|
B. callitrichidarum DSM 103152 |
Feces of emperor tamarin |
[29] |
|
B. callitrichos DSM 23973 |
Feces of common marmoset |
[26] |
|
B. canis DSM105923 |
Feces of dog |
[10] |
|
B. castoris LMG 30937 |
Feces of beaver |
[6] |
|
B. catenulatum LMG 11043 |
Adult intestine |
[30] |
|
B. catenulatum subsp. kashiwanohense DSM21854 |
Infant feces |
[31] |
|
B. catulorum DSM103154 |
Feces of common marmoset |
[32] |
|
B. cebidarum LMG31469 |
Feces of golden-headed tamarin |
[9] |
|
B. choerinum LMG 10510 |
Feces of piglet |
[28] |
|
B. commune LMG28292 |
Bumblebee gut |
[33] |
|
B. coryneforme LMG 18911 |
Hindgut of honeybee |
[25] |
|
B. criceti LMG 30188 |
Feces of European hamster |
[7] |
|
B. crudilactis LMG 23609 |
Raw cow milk |
[34] |
|
B. cuniculi LMG 10738 |
Feces of rabbit |
[28] |
|
B. dentium LMG 11045 |
Oral cavity |
[30] |
|
B. dolichotidis LMG 30941 |
Feces of Patagonian mara |
[6] |
|
B. eulemuris DSM 100216 |
Feces of black lemur |
[35] |
|
B. felsineum DSM103139 |
Feces of cotton-top tamarin |
[11] |
|
B. gallicum LMG 11596 |
Adult intestine |
[36] |
|
B. goeldii LMG 30939 |
Feces of Goeldi’s marmoset |
[6] |
|
B. hapali DSM 100202 |
Feces of baby common marmoset |
[37] |
|
B. imperatoris LMG 30297 |
Feces of emperor tamarin |
[7] |
|
B. indicum LMG 11587 |
Insect |
[25] |
|
B. italicum LMG 30187 |
Feces of European rabbit |
[7] |
|
B. jacchi DSM 103362 |
Feces of baby common marmoset |
[38] |
|
B. lemurum DSM 28807 |
Feces of ring-tailed lemur |
[39] |
|
B. leontopitechi LMG 31471 |
Feces of Goeldi’s monkey |
[9] |
|
B. longum subsp. infantis ATCC 15697 |
Intestine of infant |
[17] |
|
B. longum subsp. longum LMG 13197 |
Adult intestine |
[17] |
|
B. longum subsp. suis LMG 21814 |
Feces of pig |
[40] |
|
B. magnum LMG 11591 |
Feces of rabbit |
[30] |
|
B. margollesii LMG 30296 |
Feces of pygmy marmoset |
[7] |
|
B. meryciucm LMG 11341 |
Rumen of bovine |
[41] |
|
B. minimum LMG 11592 |
Sewage |
[42] |
|
B. mongoliense DSM 21395 |
Fermented mare’s milk |
[43] |
|
B. moukabalense DSM 27321 |
Feces of gorilla |
[44] |
|
B. myosotis DSM 100196 |
Feces of common marmoset |
[37] |
|
B. parmae LMG 30295 |
Feces of pygmy marmoset |
[7] |
|
B. platyrrhinorum SMA15 |
Feces of squirrel monkey |
[45] |
|
B. primatium DSM 100687 |
Feces of cotton-top tamarin |
[11] |
|
B. pseudocatenulatum LMG 10505 |
Infant feces |
[28] |
|
B. pseudolongum subsp. globosum LMG 11596 |
Rumen of bovine |
[46] |
|
B. pseudolongum subsp. pseudolongum LMG 11571 |
Feces of swine |
[21] |
|
B. psychraerophilum LMG 21775 |
Caecum of pig |
[47] |
|
B. pullorum subsp. gallinarum LMG 11586 |
Caecum of chicken |
[48] |
|
B. pullorum subsp. pullorum LMG 21816 |
Feces of chicken |
[8] |
|
B. ramosum DSM 100688 |
Feces of cotton-top tamarin |
[18] |
|
B. reuteri DSM 23975 |
Feces of common marmoset |
[26] |
|
B. rousetti BCRC 81136 |
Feces of Egyptian fruit bat |
[49] |
|
B. ruminantium LMG 21811 |
Rumen of bovine |
[41] |
|
B. pullorum subsp. saeculare LMG 14934 |
Feces of rabbit |
[50] |
|
B. saguini LMG 23967 |
Feces of tamarin |
[26] |
|
B. saimiriisciurei SMA1 |
Feces of squirrel monkey |
[45] |
|
B. saimirii LMG 30940 |
Feces of Bolivian saimiri |
[6] |
|
B. scaligerum DSM 103140 |
Feces of cotton-top tamarin |
[11] |
|
B. scardovii LMG 21589 |
Blood |
[51] |
|
B. simiarum DSM 103153 |
Feces of emperor tamarin |
[11] |
|
B. stellenboschense DSM 23968 |
Feces of tamarin |
[26] |
|
B. subtile LMG 11597 |
Sewage |
[42] |
|
B. porcinum LMG 21689 |
Feces of piglet |
[52] |
|
B. thermacidophilum LMG 21395 |
Anaerobic digester |
[53] |
|
B. termophilum JCM 7027 |
Rumen of bovine |
[21] |
|
B. tibiigranuli LMG 31086 |
Water kefir |
[54] |
|
B. tissieri DSM 100201 |
Feces of baby common marmoset |
[37] |
|
B. tsurumiense JCM 13495 |
Hamster dental plaque |
[55] |
|
B. vansinderenii LMG 30126 |
Feces of emperor tamarin |
[56] |
|
B. vespertilionis DSM 106025 |
Feces of Egyptian fruit bat |
[49] |
|
B. xylocopae DSM104955 |
Carpenter bee digestive tract |
[5] |
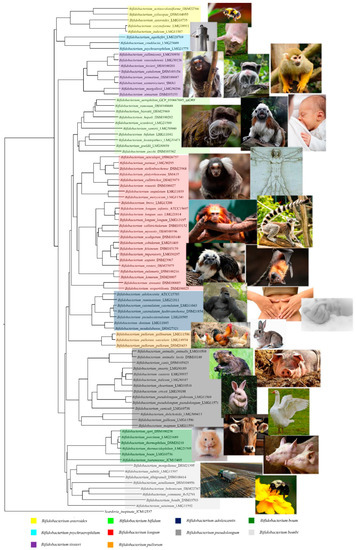
Figure 1. Phylogenetic tree of the Bifidobacterium genus based on the concatenation of 191 core amino acid sequence genes. The core genes-based tree shows the subdivision of the 10 phylogenetic groups of the Bifidobacterium genus represented with different colors. The phylogenetic tree was built by the neighbor-joining method with corresponding sequences of Scardovia inopinata JCM 12,537 being employed as outgroup. Bootstrap percentages above 50 are shown at node points, based on 100 replicates of the phylogenetic tree. The ecological origins of the various phylogenetic groups are highlighted beside the phylogenetic tree.
2. Ecology
Bifidobacteria also naturally occur in the gastrointestinal tract (GIT) of animals, such as nonhuman mammals, insects, and birds [5-11], while they have also been isolated from human blood [51], sewage [42], the oral cavity [55], and fermented milk [15]. In this context, it has been demonstrated that the ability of bifidobacteria to adapt to specific environments is species-dependent [4]. Until recently, scientific studies revealed that B. longum, B. adolescentis, B. pseudolongum, and B. bifidum species possess a cosmopolitan lifestyle [4], whereas other bifidobacterial species appear to be adapted to the GIT of particular animals (e.g., Bifidobacterium cuniculi for rabbits, Bifidobacterium angulatum for cows, and Bifidobacterium gallinarum for chickens) or the human gut (e.g., B. breve and B. longum species) [4,12]. However, recent ecological studies, based on Internally Transcribed Spacer (ITS) profiling, have revealed that the distribution of Bifidobacterium species is not host-specific [57,58]. For example, the B. breve species, which until that point had only been associated with the human gut, was shown to be present also in domesticated animals [57]. Furthermore, particular species, such as Bifidobacterium actinocoloniiforme, B. asteroides, Bifidobacterium bohemicum, B. bombi, and Bifidobacterium indicum, which were previously thought to be highly specialized to colonize the insect gut, were shown to be widely distributed among various mammalian hosts [58]. Notably, the distribution of bifidobacterial species in different ecological niches reinforces the idea that anthropogenic influences may have promoted such apparent horizontal transmission events.
The Bifidobacterium genus is one of the most abundant bacterial genera present in the human gut during the early stages of life [59-61] and these microorganisms are reported to be among the first bacterial colonizers of the newborn’s GIT [62]. It has been demonstrated that bifidobacteria may engage in vertical transmission that occurs between a mother and her newborn during birth and possibly through subsequent breastfeeding [63,64]. This fascinating phenomenon not only occurs in human beings [65] but also in other mammalian species [58]. In this context, some studies have shown how taxonomic classification of bifidobacteria present in the mother’s microbiota strongly correlates with that of the infant [66,67]. In particular, a study based on ITS-profiling and shotgun-metagenomics approaches has led to the identification of the species shared between a mother and her child [68]. In this study, the microbiota of a mother’s fecal and milk samples were assayed together with corresponding infant fecal samples collected at different time points. These analyses demonstrated that in some cases, identical bifidobacterial strains are shared in both mother’s and baby’s gut microbiota [68]. A B. breve strain and B. longum subsp. longum isolate were seen to be among the protagonists of vertical transmission from mother to child, being found both in the newborn’s meconium and in the fecal samples of the child for up to 90 days [68]. Several species of this genus are believed to have undergone specific genetic and metabolic adaptations in order to facilitate colonization of the infant gut, for example, the ability of certain bifidobacterial species and strains to metabolize specific oligosaccharides present in human milk [69]. Specifically, bifidobacterial species that are prevalent in the gut of infants include B. breve, B. longum subsp. infantis, B. longum subsp. longum, B. pseudocatenulatum, and B. bifidum [62], whereas B. adolescentis, B. catenulatum, B. pseudocatenulatum, and B. longum subsp. longum [70,71] are commonly occurring species in the adult intestine. In this context, it is not fully correct to consider the use of fecal material as a representation of the entire intestinal microbiota. In fact, the fecal microbiota consists not only of mucosal adherent members of the human GIT microbiota but also of transient bacteria derived from the diet or other environmental microbial contaminations [72]. Specifically, only a small number of bifidobacterial species (i.e., B. longum, B. adolescentis, B. breve, B. pseudocatenulatum, and B. pseudolongum) seem to be dominant in the examined biopsies, whereas certain other bifidobacterial species are restricted to a specific ecological niche (e.g., B. bifidum and B. pseudolongum) [73]. Analyses not only of human intestinal mucosal but also of fecal samples have shown that bifidobacterial distribution changes within ages, with a remarkable conservation in terms of species and strains in adults and children [73]. Furthermore, little is known about the diversity of bifidobacterial populations occurring between individuals and between different compartments of the GIT within the same individual [73].
The presence of bifidobacteria in the GIT has been associated with various health benefits, including the development of the immune system, protection against pathogens mediated through the process of competitive exclusion, and/or the production of metabolites such as short-chain fatty acids (SCFA) and vitamins [59,62,70,74]. Indeed, human-residential bifidobacteria (HRB) are also capable of producing folate, also known as vitamin B9 or B11, which is required for an efficient DNA replication, DNA repair/methylation, and synthesis of nucleotides, vitamins, and certain amino acids [75,76]. For these reasons, several bifidobacterial strains/species are used as active ingredients in a variety of so-called functional foods due to their perceived health-promoting or probiotic properties [2]. In this context, probiotic bifidobacterial strains belonging to B. longum and Bifidobacterium animalis subsp. lactis species are usually added to yogurt, other fermented milks, and, more recently, to cheese, which are the most popular probiotic foodstuffs at the moment [77,78]. Moreover, clinical studies have demonstrated that B. animalis ssp. lactis Bb-12, administered as probiotic adjunctive therapy, have beneficial effects in the case of infectious diarrhea caused by viruses or bacteria [79,80], decreasing the frequency or shortening the duration of the infection and increasing immune responses [81].
This entry is adapted from the peer-reviewed paper 10.3390/microorganisms9010008