The adrenal gland is a complex endocrine organ composed of two components: a steroidogenic tissue, which produces steroid hormones, and a chromaffin tissue, which mainly produces norepinephrine and epinephrine. Through evolution, their relationships with each other changed. They begin as isolated chromaffin and steroidogenic cell aggregates, typical of fish, and end with the advanced compact gland, typical of mammals, which consists of an external steroidogenic cortical zone and an internal chromaffin medullary zone. The adrenal gland of reptiles is unique because, with few exceptions, it is near the gonads and genital ducts, and the chromaffin and steroidogenic tissues are closely associated.
- adrenal gland
- chromaffin tissue
- NE/E cell ratio
- Reptilia
- Squamata
- steroidogenic tissue
1. The Adrenal Gland
2. Paracrine Relationships in the Adrenal Gland
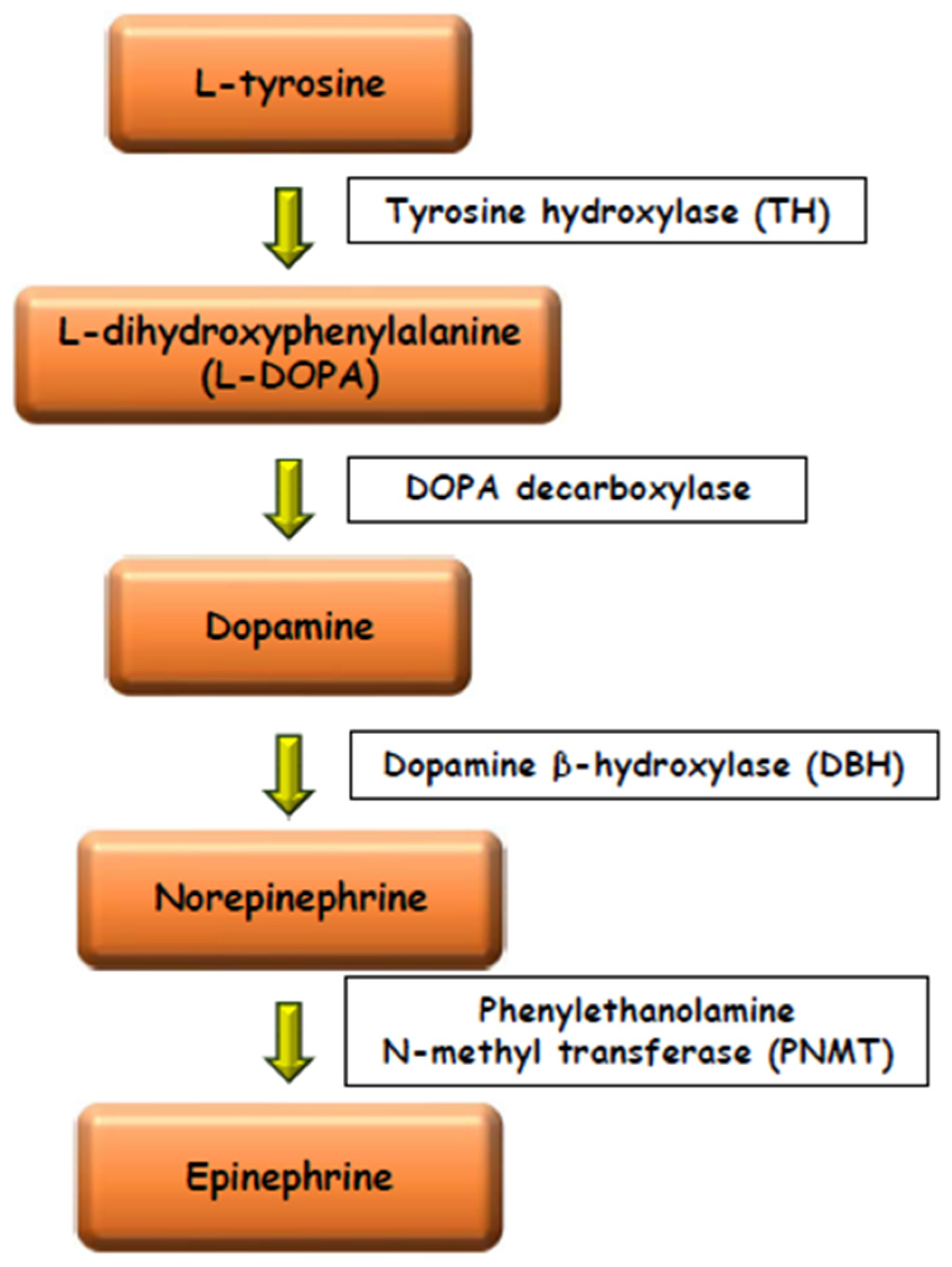
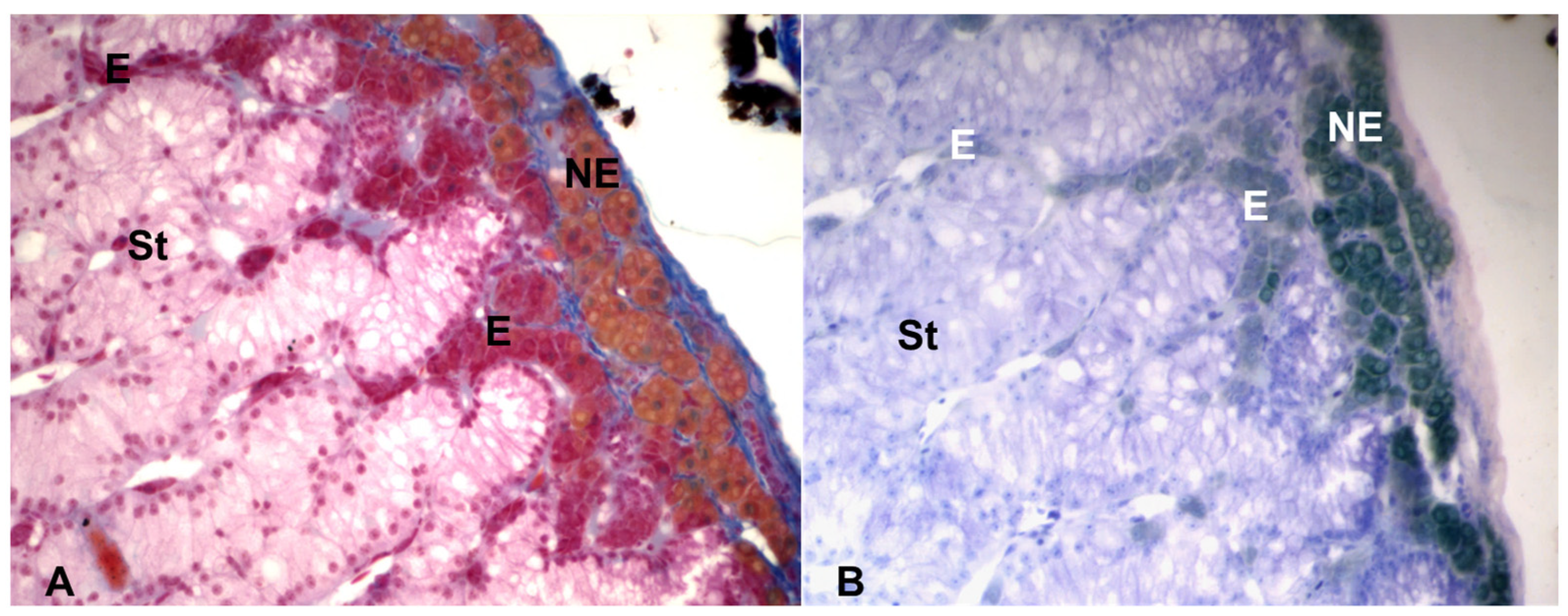
3. The Morphology of the Chromaffin and the Steroidogenic Tissues
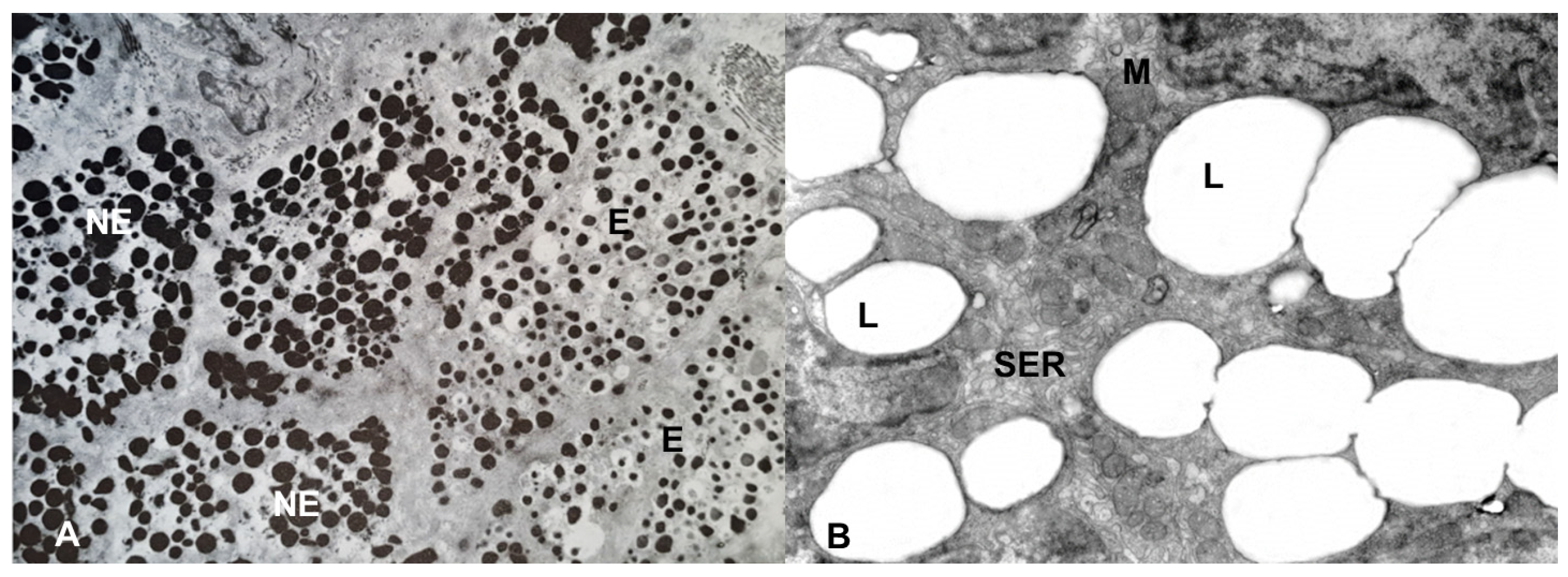
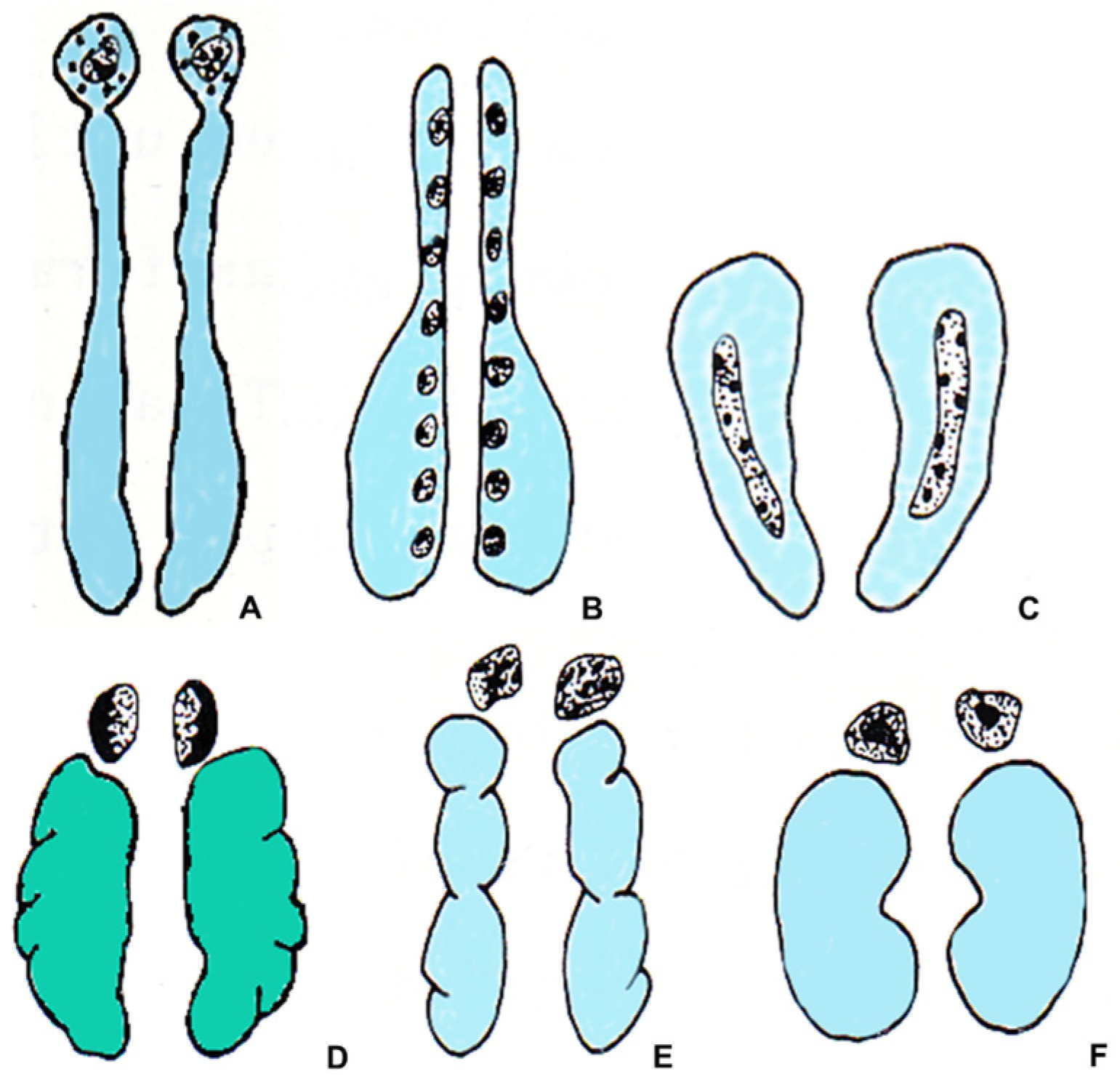
4. The NE/E Cell Ratio
5. The Adrenal Gland of Reptiles
6. The Adrenal Gland of Squamata
This entry is adapted from the peer-reviewed paper 10.3390/ani13172686
References
- Perry, F.S.; Capaldo, A. The autonomic nervous system and chromaffin tissue: Neuroendocrine regulation of catecholamine secretion in non-mammalian vertebrates. Auton. Neurosci. Basic Clin. 2011, 165, 54–66.
- Varano, L.; Laforgia, V. Evolutionary trends in the adrenal gland of reptiles. In Symposium on the Evolution of Terrestrial Vertebrates; Ghiara, G., Angelini, F., Olmo, E., Varano, L., Eds.; Mucchi: Modena, Italy, 1991; Volume 4, pp. 291–303.
- Norris, D.O. Vertebrate Endocrinology, 4th ed.; Academic Press: Amsterdam, The Netherlands, 2007.
- Accordi, F. The chromaffin cells of urodele amphibians. J. Anat. 1991, 179, 1–8.
- Grassi Milano, E.; Accordi, F. Evolutionary trends in adrenal gland of anurans and urodeles. J. Morphol. 1986, 189, 249–259.
- Grassi Milano, E. Development of the adrenal gland in amniotes: A comparison between chelonians and birds. Boll. Zool. 1991, 58, 205–209.
- Di Lorenzo, M.; Barra, T.; Rosati, L.; Valiante, S.; Capaldo, A.; De Falco, M.; Laforgia, V. Adrenal gland response to endocrine disrupting chemicals in fishes, amphibians and reptiles: A comparative overview. Gen. Comp. Endocrinol. 2020, 297, 113550.
- Carsia, R.V.; McIlroy, P.J.; John-Alder, H.B. Adrenocortical function in avian and non-avian reptiles: Insights from dispersed adrenocortical cells. Comp. Biochem. Physiol. A 2023, 281, 111424.
- Ehrart-Bornstein, M.; Hinson, J.P.; Bornstein, S.R.; Scherbaum, W.A.; Vinson, G.P. Intraadrenal interactions in the regulation of adrenocortical steroidogenesis. Endocr. Rev. 1998, 19, 101–143.
- Rao, F.; Zhang, L.; Wessel, J.; Zhang, K.; Wen, G.; Kennedy, B.P.; Rana, B.K.; Das, M.; Rodriguez-Flores, J.L.; Smith, D.W.; et al. Tyrosine hydroxylase, the rate-limiting enzyme in catecholamine biosynthesis: Discovery of common human genetic variants governing transcription, autonomic activity, and blood pressure in vivo. Circulation 2007, 116, 993–1006.
- Lucas, C.A.; Thoenen, H. Selective induction by glucocorticoids of tyrosine hydroxylase in organ cultures of rat pheochromocytoma. Neuroscience 1977, 2, 1095–1101.
- Kelner, K.L.; Pollard, H.B. Glucocorticoid receptors and regulation of phenylethanolamine-N- methyltransferase activity in cultured chromaffin cells. J. Neurosci. 1985, 5, 2161–2168.
- Evinger, M.J.; Towle, A.C.; Park, D.H.; Lee, P.; Joh, T.H. Glucocorticoids stimulate transcription of the rat phenylethanolamine N-methyltransferase (PNMT) gene in vivo and in vitro. Cell. Mol. Neurobiol. 1992, 12, 193–215.
- Ghosh, A.; Carmichael, S.W.; Mukherjee, M. Avian adrenal medulla: Cytomorphology and function. Acta Biol. Szeged. 2001, 45, 1–11.
- Hodel, A. Effects of glucocorticoids on adrenal chromaffin cells. J. Neuroendocr. 2001, 13, 216–220.
- Wurtman, R.J. Control of epinephrine synthesis in the adrenal medulla by the adrenal cortex: Hormonal specificity and dose-response characteristics. Endocrinology 1966, 79, 392–394.
- Unsicker, K.; Huber, K.; Schober, A.; Kalcheim, C. Resolved and open issues in chromaffin cell development. Mech. Dev. 2013, 130, 324–329.
- Zachariasen, R.D.; Newcomer, W.S. Phenylethanolamine-N-methyltransferase activity in the avian adrenal following immobilization or adrenocorticotropin. Gen. Comp. Endocrinol. 1974, 23, 193–198.
- Laforgia, V.; Varano, L. The influence of the interrenal steroidogenic tissue on the chromaffin cells of the adrenal gland of Lacerta s. sicula Raf.: Effects of ACTH administration during the winter. Cell. Mol. Biol. 1978, 23, 379–390.
- Capaldo, A.; Laforgia, V.; Sciarrillo, R.; De Falco, M.; Valiante, S.; Gay, F.; Virgilio, F.; Varano, L. Effects of dopamine on the adrenal gland of Podarcis sicula (Reptilia, Lacertidae). Gen. Comp. Endocrinol. 2004, 135, 17–24.
- Capaldo, A.; Sciarrillo, R.; Valiante, S.; Gay, F.; Virgilio, F.; Varlese, M.G.; Laforgia, V.; Varano, L. Neuropeptide Y (NPY) modulates pituitary-adrenal axis activity in the lizard, Podarcis sicula. Gen. Comp. Endocrinol. 2004, 137, 237–247.
- Reid, S.G.; Vijayan, M.M.; Perry, S.F. Modulation of catecholamine storage and release by the pituitary-interrenal axis in the rainbow trout, Oncorhynchus Mykiss. J. Comp. Physiol. B 1996, 165, 665–676.
- Capaldo, A.; Gay, F.; De Falco, M.; Virgilio, F.; Laforgia, V.; Varano, L. The adrenal gland of newt Triturus carnifex (Amphibia, Urodela) following in vivo betamethasone administration. Anat. Embriol. 2006, 211, 577–584.
- Ehrhart-Bornstein, M.; Bornstein, S.R. Cross-talk between adrenal medulla and adrenal cortex in stress. Ann. N. Y. Acad. Sci. 2008, 1148, 112–117.
- Hanke, W. Changes of the biology of the adrenal cortex in the Vertebrate evolution. Nova Acta Leopold. NF 1984, 56, 363–378.
- Laforgia, V.; Capaldo, A. Annual cycle of the chromaffin cells of Triturus cristatus. J. Morphol. 1991, 208, 83–90.
- Gay, F.; Valiante, S.; Sciarrillo, R.; De Falco, M.; Laforgia, V.; Capaldo, A. Annual and daily serum aldosterone and catecholamine patterns in males of the Italian crested newt, Triturus carnifex (Amphibia, Urodela). Ital. J. Zool. 2010, 77, 384–390.
- Gay, F.; Laforgia, V.; Capaldo, A. Human follicle-stimulating hormone modulation of adrenal gland activity in the Italian crested newt, Triturus carnifex (Amphibia, Urodela). Comp. Biochem. Physiol. A-Mol. Int. Physiol. 2008, 151, 126–132.
- Coupland, R.E. The natural history of the chromaffin cell. Twenty-five years on the beginning. Arch. Histol. Cytol. 1989, 52, 331–341.
- Giordano-Lanza, G. Sulla istofisiologia degli elementi cellulari della parte midollare del surrene. Quad. Anat. Prat. S 1961, 17, 251–284.
- Brown, W.J.; Barajas, L.; Latta, H. The ultrastructure of the human adrenal medulla with comparative studies of white rat. Anat. Rec. 1971, 169, 173–184.
- Khan, M.B.; Lee, B.R.; Kamitani, T. A simple and sensitive method for the demonstration of norepinephrine-storing adrenomedullary chromaffin cells. Histochem. Cell Biol. 2012, 138, 155–165.
- Wood, J.G. Identification of and observations on epinephrine and norepinephrine containing cells in the adrenal medulla. Am. J. Anat. 1963, 112, 285–304.
- Pearse, A.G.E. Histochemistry Theoretical and Applied; Little, Brown & Co.: Boston, MA, USA, 1960.
- Tramezzani, J.H.; Chiocchio, S.; Wasserman, G.F. A technique for light and electron microscopic identification of adrenaline and noradrenaline storing cells. Cytochemistry 1964, 12, 890–899.
- Capaldo, A.; Laforgia, V.; Sciarrillo, R.; Valiante, S.; Gay, F.; Varano, L. Localization and role of serotonin in the adrenal gland of Podarcis sicula (Reptilia, Lacertidae). Gen. Comp. Endocrinol. 2003, 132, 66–76.
- Capaldo, A.; De Falco, M.; Rosati, L.; Laforgia, V. Transmission electron microscopy: A method for studying the adrenal chromaffin cells. In Chromaffin Cells. Methods in Molecular Biology; Borges, R., Ed.; Humana: New York, NY, USA, 2023; Volume 2565, pp. 43–55.
- Gonzàlez-Santana, A.; Castañeira, L.; Baz-Dàvila, R.; Estévez-Herrera, J.; Domìnguez, N.; Méndez-Lòpez, I.; Padìn, J.F.; Castañeira, A.; Machado, J.D.; Ebert, S.N.; et al. Adrenergic chromaffin cells are adrenergic even in the absence of epinephrine. J. Neurochem. 2020, 152, 299–314.
- Moawad, U.K.; Soliman, S.M.M.; Mazher, K.M.; Hassan, R.M.; Nabil, T.M. Histological, histochemical, ultrastructural and immunohistochemical identification and characterization of the neurosecretory cells of the adult rabbit’s adrenal medulla. Anat. Histol. Embryol. 2022, 51, 280–288.
- Maneu, V.; Borges, R.; Gandìa, L.; Garcìa, A.G. Forty years of the adrenal chromaffin cell through ISCCB meetings around the world. Pflug. Arch. Eur. J. Physiol. 2023, 475, 667–690.
- Idelman, S. The structure of the mammalian adrenal cortex. In General, Comparative and Clinical Endocrinology of the Adrenal Cortex; Chester Jones, I., Henderson, I.W., Eds.; Academic Press: London, UK; New York, NY, USA; San Francisco, CA, USA, 1978; pp. 1–199.
- Klingbeil, C.K.; Holmes, W.N.; Pearce, R.B.; Cronshaw, J. Functional significance of interrenal cell zonation in the adrenal gland of the duck (Anas platyrhynchos). Cell Tissue Res. 1979, 201, 23–36.
- Holmes, W.N.; Cronshaw, J. Adrenal gland: Some evidence for the structural and functional zonation of the steroidogenic tissues. J. Exper. Zool. 1984, 232, 627–631.
- Lofts, B.; Phillips, J.G. Some aspects of the structure of the adrenal gland in snakes. J. Endocrinol. 1965, 33, 327–328.
- Mitani, F. Functional zonation of the rat adrenal cortex: The development and maintenance. Proc. Jpn. Acad. Ser. B 2016, 90, 163–183.
- Vinson, G.P. Functional Zonation of the Adult Mammalian Adrenal Cortex. Front. Neurosci. 2016, 10, 238.
- Ghosh, A. Cytophysiology of the avian adrenal medulla. In International Review of Cytology; Bourne, G., Danielli, J.P., Eds.; Academic Press: New York, NY, USA, 1977; pp. 253–284.
- Braun, E.L.; Kimball, R.T. Data types and the phylogeny of Neoaves. Birds 2021, 2, 1–22.
- Arnold, E.N. Relationships of the paleartctic lizards assigned to the genera Lacerta, Algyroides and Psammodromus (Reptilia: Lacertidae). Bull. Br. Mus. (Nat. Hist.) Zool. 1973, 25, 291–366.
- Lutz, B.; Cei, J.M. Immunological data on the taxonomy of some Italian lizards (Reptilia, Lacertidae). Mon. Zool. Ital. 1977, 11, 231–236.
- Lanza, B.; Cei, J.M.; Crespo, E.G. Immunological investigations on the taxonomic status of some Mediterranean lizards (Reptilia, Lacertidae). Mon. Zool. Ital. 1977, 11, 211–221.
- Lutz, D.; Mayer, W. Albumin evolution and its phylogenetic and taxonomic implications in several lacertid lizards. Amphibia-Reptilia 1985, 6, 53–61.
- Conrad, J.L. Phylogeny and systematics of Squamata (Reptilia) based on morphology. Bull. Am. Mus. Nat. Hist. 2008, 310, 1–182.
- Mulcahy, D.G.; Noonan, B.P.; Moss, T.; Townsend, T.M.; Reeder, T.W.; Sites, J.W., Jr.; Wiens, J.J. Estimating divergence times and evaluating dating methods using phylogenomic and mitochondrial data in squamate reptiles. Mol. Phylogenet Evol. 2012, 65, 974–991.
- Townsend, T.M.; Larson, A.; Louis, E.; Macey, J.R. Molecular phylogenetics of Squamata: The position of snakes, amphisbaeninans, and dibamids, and the root of the squamate tree. Syst. Biol. 2004, 53, 735–757.
- Vidal, N.; Hedges, S.B. The phylogeny of squamate reptiles (lizards, snakes, and amphisbaenians) inferred from nine nuclear protein-coding genes. CR Biol. 2005, 328, 1000–1008.
- Wiens, J.J.; Hutter, C.R.; Mulcahy, D.G.; Noonan, B.P.; Townsend, T.M.; Sites, J.W.; Reeder, T.W. Resolving the phylogeny of lizard and snakes (Squamata) with extensive sampling of genes and species. Biol. Lett. 2012, 8, 1043–1046.
- Pyron, R.A.; Burbrink, F.T.; Wiens, J.J. A phylogeny and revised classification of Squamata, including 4161 species of lizards and snakes. BMC Evol. Biol. 2013, 13, 93.
- Modesto, S.P.; Anderson, J.S. The phylogenetic definition of Reptilia. Syst. Biol. 2004, 53, 815–821.
- The NCBI Taxonomy Database. Available online: https://www.ncbi.nlm.nih.gov/Taxonomy/Browser/wwwtax.cgi (accessed on 30 June 2023).
- Gabe, M.; Martoja, M. Contribution à l’histologie de la glande surrénale des Squamata. Archs Anat. Microsc. 1961, 50, 1.
- Gabe, M.; Rancurel, P. Contribution à l’histologie de la glande surrénale de Crocodylus niloticus Laur. Archs Anat. Microsc. 1964, 53, 225–240.
- Gabe, M.; Saint Girons, H. Particularités histologiques de la glande surrénale chez Sphenodon punctatus Gray. C. R. Hebd. SÉAnc Acad. Sci. Paris 1964, 258, 3559–3562.
- Wasserman, G.F.; Tramezzani, J.H. Separate distribution of adrenaline and noradrenaline secreting cells in the adrenal of snakes. Gen. Comp. Endocrinol. 1963, 3, 480–489.
- Varano, L.; Della Corte, F.; Galgano, M. Brevi note sulla ultrastruttura delle cellule a catecolamine di Lacerta s. sicula Raf. Atti Soc. Pel. Sci. Fis. Mat. Nat. 1969, 15, 39–44.