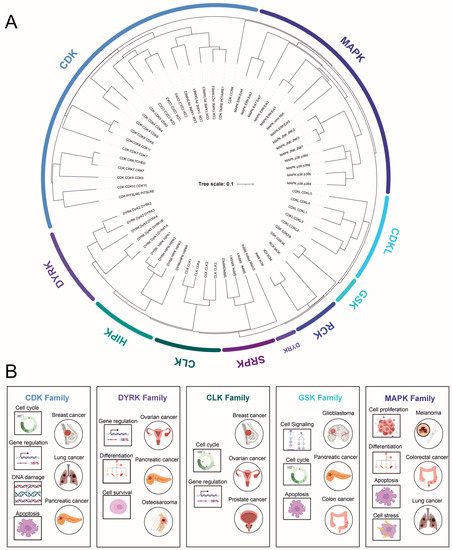
2.2. Mitogen-Activated Protein Kinases (MAPKs)
The mitogen-activated protein kinase (MAPK) family consists of 14 serine/threonine kinases that can be divided into atypical and conventional kinases.
The atypical mitogen-activated protein kinases (MAPKs) form a distinct, unique subgroup within the larger MAPK family, a major regulator of cellular processes. This group includes extracellular signal-regulated kinase 3 (ERK3), ERK4, ERK7, and Nemo-like kinase (NLK) [
46]. Unlike their typical counterparts, atypical MAPKs feature a divergence from the canonical MAPK architecture in their kinase domain, deviating from the conserved TXY motif crucial for activation [
47]. Moreover, they exhibit unique activation and regulatory mechanisms, such as autophosphorylation [
48]. Their specific physiological roles and functions remain more undefined compared to classical MAPKs. However, emerging evidence links atypical MAPKs to several critical cellular processes, including cell differentiation, proliferation, motility, and apoptosis [
49]. For instance, ERK3 and ERK4 are associated with cell migration and actin cytoskeleton reorganization, while ERK7/8 plays roles in cell cycle progression and response to stress [
50]. In spite of this, the mechanisms by which atypical MAPKs exert their influence and their potential implications in human diseases, such as cancer, are areas of ongoing research and discovery.
The conventional kinases can be further subdivided into four subfamilies: (1) extracellular signal-regulated kinases 1/2 (ERK1/2), (2) c-Jun amino-terminal kinases 1, 2, and 3 (JNK1-3), (3) p38 MAPKs, and (4) extracellular signal-regulated kinase 5 (ERK5). These enzymes are characterized by a system in which three kinases act sequentially to phosphorylate the downstream kinase. Activators for ERKs are MEK1 and MEK2; for JNKs-, MKK4 and MKK7; for p38-, MKK3 and MKK6; and for ERK5-, MEK5. MAPKs are highly conserved, respond to extracellular signaling cues, and regulate numerous essential cellular processes such as proliferation, differentiation, stress responses, and the transcriptional control of CDKs in organisms ranging from yeast to humans. Over the past few decades, extensive research has been conducted on MAPKs, from their substrates and functions to their roles in health and cancer.
Extracellular signal-regulated kinases 1/2 respond to various stimuli, including growth factors, insulin, cytokines, and carcinogens [
50,
51]. They are components of the cell surface receptor (receptor tyrosine kinase)/RasGTP/Raf/MEK/ERK signaling cascade. Components of this signaling cascade are essential for proper cell proliferation. ERKs function both in the cytoplasm to promote cell proliferation (G1 to S phase transition) and in the nucleus to phosphorylate and activate numerous transcription factors (TFs) [
50,
52]. Upon TF activation, c-Jun is expressed and stabilized by direct phosphorylation by ERK1/2 [
53], allowing association with c-Fos and the subsequent formation of the AP-1 complex. AP-1 is essential for cyclin D1 expression, which interacts with CDKs to drive G1 to S transition.
-
c-Jun N-terminal kinases are highly homologous (>85%) but have a distinct tissue distribution. JNK1 and 2 are ubiquitously expressed, while JNK3 expression is mainly limited to the brain. In contrast to ERKs, JNKs are primarily activated by stress signals such as oxidative stress, radiation, and DNA-damaging agents. JNKs are mainly localized in the cytoplasm, but their identified substrates are mostly TFs, including c-Jun, p53, STAT3, and c-Myc. The phosphorylation of c-Jun leads to AP-1 complex formation and thus the transcription of cyclin D1, promoting cell cycle progression, similar to ERK1/2 [
54]. To date, only a few cytoplasmic interaction partners of JNKs have been identified [
50,
55].
-
The p38 subfamily consists of four members (α, β, γ, δ), which respond to various environmental stress stimuli and cytokines such as interleukin-1 and tumor necrosis factor α(TNF). Interestingly, p38 both regulates the production of cytokines and responds to them. Other targets of p38 regulation are TFs and other protein kinases. Based on observations of p38 activation, it plays a role in inflammation, cell cycle regulation, and apoptosis [
50,
56].
-
ERK5 (BMK1 or big MAP kinase 1) has a kinase domain similar to ERK1/2, sharing 51% similarity with ERK2. ERK5 is essential during normal embryogenesis [
57]. An upstream activator of ERK5 is MEK5, whose expression is elevated in metastatic prostate cancer [
58]. Similar to ERK1/2 and JNK, ERK5 also promotes cyclin D1 expression and cell cycle progression [
59], as well as plays a crucial role in the maintenance of mitochondrial function and neuronal survival [
60]. ERK5 is involved in various cellular processes, including cell survival, differentiation, and angiogenesis. Its activation has been linked to growth factors, oxidative stress, and other extracellular stimuli.
-
The MAPK pathway has a critical role in cancer biology, extending beyond the extensively studied BRAF mutation in melanoma. The MAPK/ERK pathway, for instance, has been implicated in colorectal cancer, with mutations in KRAS and NRAS genes leading to its persistent activation, promoting uncontrolled cell proliferation and tumor growth [
61]. These mutations, unfortunately, render the tumors resistant to EGFR-targeted therapies, highlighting the need for novel therapeutic strategies [
62].
-
Additionally, the JNK MAPK pathway, associated primarily with responses to stress signals and apoptosis, has shown links to cancer biology. Aberrations in JNK signaling can lead to an imbalance between cell proliferation and death, thereby contributing to oncogenesis. For instance, overactive JNK signaling has been found in several cancers, including breast and gastric cancer, often correlating with a worse prognosis [
63]. Furthermore, the p38 MAPK pathway, typically associated with inflammation and cell differentiation, is also relevant in cancer research. Its complex, dual role in tumorigenesis is being unraveled; while its activation can suppress tumor growth by promoting cell cycle arrest and apoptosis, chronic activation can also enhance cancer cell survival, contributing to chemoresistance [
64].
In summary, the MAPK family comprises a diverse group of kinases that play crucial roles in cellular signaling pathways. These kinases are involved in various cellular processes, including proliferation, differentiation, and stress responses, and are integral to the transcriptional control of CDKs. Alterations in the function of the components of the cellular signaling cascade can lead to cancer. Thus, understanding the mechanisms and functions of MAPKs is essential for advancing our knowledge of cellular processes and uncovering potential therapeutic targets in various diseases.
2.3. Glycogen Synthase Kinase-3 (GSK-3)
Glycogen synthase kinase-3 is a conserved, ubiquitously expressed serine/threonine kinase with unique characteristics, such as its high activity in resting cells. Humans have two forms of GSK-3s: GSK-3α and GSK-3β, which share an 85% overall sequence homology [
65]. The kinase domain of these kinases is highly homologous (98%), while the N- and C-terminal domains differ [
65]. GSK-3α and GSK-3β have distinct functions, as evidenced by the fact that homozygous GSK-3β knockout mice are embryonic-lethal, whereas GSK3α knockout mice are viable [
66,
67].
Unlike the CDK and MAPK families, GSK activity is regulated through multiple levels and mechanisms, ranging from phosphorylation by other kinases, autophosphorylation, and priming phosphorylation of the target substrate by another kinase [
68,
69,
70]. In most cases, the phosphorylation of serine residues inhibits GSK’s kinase activity, while tyrosine phosphorylation increases its activity [
71]. GSK-3 (mainly GSK-3α) has been extensively studied and is known to function in various signaling pathways, including Wnt, Notch, and Hedgehog (proliferation), as well as growth factors affecting differentiation/survival. It has been implicated in multiple roles in many human pathological conditions, ranging from neurodegenerative diseases to diabetes and several types of cancers [
70,
72].
GSK is a critical component of the canonical Wnt signaling pathway, which is essential during normal development and is often dysregulated in cancers. The main events during Wnt-β-catenin signaling involve a central component known as the destruction complex that regulates cytoplasmic β-catenin levels. This complex comprises tumor suppressors Axin and adenomatosis polyposis coli (APC), as well as casein kinase-1α (CK1α) and GSK-3 [
73]. In the absence of the Wnt-ligand, β-catenin is assembled into this complex and phosphorylated by CK1 and GSK-3, marking it for proteasomal degradation and thus preventing the transcription of β-catenin-dependent genes. When Wnt binds to its receptor Frizzled, the destruction complex cannot phosphorylate and ubiquitinate β-catenin, allowing it to translocate to the nucleus, where it forms complexes with transcription factors and promotes transcription. Phosphorylation by GSK-3 is also essential for the targeting of multiple other proteins for proteasomal degradation [
72,
74]. In addition to its role in Wnt signaling, GSK-3 has been implicated in other critical signaling pathways. For example, GSK-3 plays a role in the insulin signaling pathway, which is essential for glucose homeostasis. Abnormalities in GSK-3 function have been linked to insulin resistance and the development of type 2 diabetes [
75]. GSK-3 inhibitors have shown promise in preclinical studies as potential therapeutic agents for the treatment of type 2 diabetes [
76].
GSK-3 has been found to be overactive in various types of cancers, including colon, breast, and prostate cancer. The overactivation of GSK-3 can lead to increased cell proliferation, migration, and invasion, as well as reduced apoptosis [
70]. Targeting GSK-3 may represent a potential therapeutic strategy for cancer treatment. Preclinical studies have shown that GSK-3 inhibitors can suppress tumor growth and metastasis, as well as sensitize cancer cells to chemotherapy and radiation therapy [
72].
In summary, GSK-3 is a multifunctional kinase that plays a pivotal role in numerous signaling pathways and cellular processes. The dysregulation of GSK-3 activity has been implicated in various pathological conditions. Consequently, GSK-3 has emerged as a potential therapeutic target, and the development of GSK-3 inhibitors may hold promise for the treatment of these diseases. However, due to the pleiotropic nature of GSK-3, further research is needed to understand the precise mechanisms of GSK-3 regulation and its involvement in different pathologies to develop safe and effective therapeutic strategies.
2.4. Dual-Specificity Tyrosine (Y)-Phosphorylation-Regulated Kinases (DYRKs)
DYRK is a relatively large, mammalian, dual-specificity protein kinase family that, based on homology analysis, consists of three subfamilies with a total of 10 members: DYRK (DYRK1A-B, 2–4), HIPKs (homeodomain-interacting protein kinase 1–4), and PRP4s (pre-mRNA processing protein 4 kinase).
2.5. Cdc2-like Kinase (CLK) and Other Less-Studied Kinases
The Cdc2-like kinase (CLK) family consists of four dual-specificity kinases (CLK1–4) that autophosphorylate on tyrosine residues and specifically phosphorylate serine/threonine residues on their substrates [
97]. CLKs are found in both the cytoplasm and the nucleus, where they phosphorylate the arginine- and serine-rich (RS) domains of serine/arginine (SR) splicing factor proteins, thereby controlling their nuclear distribution [
98]. SR splicing factor proteins are extensively phosphorylated by multiple kinases, and the proper phosphorylation status has been shown to be essential for SR protein activity [
99]. These proteins play a crucial role in alternative mRNA splicing, a fundamental process responsible for generating the complex human proteome during normal development.
In a recent study, CLKs were identified as upstream regulators of Aurora B, acting as novel components in the final steps of cytokinesis, the division of the cytoplasm into two daughter cells [
100]. However, as Petsalaki and Zachos concluded, our current understanding of CLKs’ cellular functions remains limited [
100]. A 2017 study reported that CLK inhibition could modulate the alternative splicing of Mcl-1, an anti-apoptotic protein, and promote cancer cell death, suggesting a potential therapeutic approach for cancer treatment [
101]. Given the importance of alternative mRNA splicing in the regulation of gene expression and the emerging roles of CLKs in cell division, further investigation into the functions and regulation of CLKs could provide valuable insights into their role in various cellular processes and their potential as therapeutic targets in human diseases.
2.6. SR-Specific Protein Kinase (SRPK)
Serine–arginine protein kinases (SRPKs) specifically phosphorylate the serine residues in serine/arginine (SR) dipeptides. They are primarily localized in the cytoplasm, where these constitutively active kinases regiospecifically phosphorylate SR splicing factors on multiple serines, inducing the translocation of these factors into the nucleus. In the nucleus, SR splicing factors are further phosphorylated by CLKs [
102,
103]. The change in the cellular localization of SR proteins is essential for phosphorylation by CLKs [
98]. As mentioned earlier, SR protein phosphorylation is necessary for correct mRNA splicing and maturation.
Over 100 SR-domain-containing proteins have been identified in the human genome [
104], indicating that there are many open questions regarding the functional roles of SRPKs in mammalian cells. SRPK1 expression is elevated in breast, colon, and pancreatic cancers [
105], as well as in acute T-cell leukemia [
106]. A 2020 study identified SRPK1 as a potential therapeutic target in glioblastoma, with the SRPK1 inhibitor SPHINX31 showing promising results in preclinical models [
107]. SRPK2 has a specific role in phosphorylating apoptosis-promoting protein ACIN1 and increasing cyclin A1 expression in leukemia cells [
108]. Given the importance of SRPKs in the regulation of alternative splicing and their association with various types of cancer, further studies on the functional roles of these kinases and their potential as therapeutic targets in human cancer are warranted.
2.7. Tyrosine Kinase Gene v-Ros Cross-Hybridizing Kinase (RCK)
The RCK serine/threonine-protein kinase family consists of three kinases: MAK, ICK, and MOK. The functions of these kinases are poorly understood. Structurally, the RCK family resembles both MAPKs and CDKs [
5]. MAK and ICK autophosphorylate on a tyrosine residue, but they need a second phosphorylation on their MAPK-like motif in their activation loop by an upstream kinase for full enzymatic activity [
109,
110,
111]. Different from MAPK, the activating kinase for MAK and ICK is cell-cycle-related kinase (CCRK), but for MOK, it remains unknown [
111,
112].
-
MAK (male germ cell-associated kinase) is mainly expressed in testicular germ cells during spermatogenesis and in the retina. In the retina, MAK localizes to connecting cilia in photoreceptor cells, negatively regulates the length of their cilia, and is essential for the survival of these cells [
113]. Not surprisingly, MAK mutations are associated with retinitis pigmentosa, a photoreceptor degeneration disease in the retina [
114,
115].
-
ICK (intestinal cell kinase) is highly conserved, constitutively, and widely expressed. Similarly to MAK, it negatively regulates ciliary length and is identified as an essential component of sonic hedgehog signaling [
116]. These two factors seem to be the underlying cause of human ECO syndrome, a multi-organ illness affecting the endocrine, cerebral, and skeletal systems, caused by a missense mutation in the ICK gene [
117]. In 2017, a study reported the role of ICK in colorectal cancer progression and its potential as a therapeutic target for the treatment of colorectal cancer [
118].
The cellular function of the third RCK, MOK (MAPK/MAK/MRK-overlapping kinase), is largely unknown. To date, only the expression of MOK in mouse wild-type and cancerous intestinal cells has been analyzed by Western blotting, showing the downregulation of MOK in adenomas [
80]. This observation suggests that MOK might have a role in intestinal cancer development, but further studies are needed.
2.8. Cyclin-Dependent Kinase-like (CDKL)
This family of five members of serine/threonine kinases with homology to MAPK and CDK families is relatively uncharacterized. Studies have focused on CDKL5, a ubiquitous protein mainly expressed in the brain, testes, and thymus [
119]. In the context of cancer, CDKL proteins are now emerging as potential players. Studies suggest that the dysregulation of CDKL5 may contribute to tumorigenesis in certain cancers. For instance, the aberrant expression of CDKL5 has been suggested in glioma and some evidence exists on its role in breast cancer, indicating its possible role in disease progression and patient prognosis [
120].
Similarly, another family member, CDKL1, has been associated with colorectal cancer, with its overexpression correlating with poor survival outcomes [
121]. These preliminary findings suggest that CDKL proteins could potentially act as novel markers for cancer diagnosis or targets for therapeutic intervention.
3. Protein Kinase Therapeutics—Kinase Inhibitors
The protein kinase domain is among the most commonly encountered domain in known cancer genes [
122]. As described earlier, most of the CMGC kinases affect various cellular signaling pathways that control critical processes like cell cycle progression, proliferation, differentiation, apoptosis, or survival. Often, abnormal phosphorylation is either the cause or consequence in cancer [
123]. Therefore, protein kinases are among the most studied druggable targets in pharmacological research [
124,
125]. In human cancers, kinases are often found overexpressed or overactive, due to a vast array of genetic and epigenetic events, including point mutations, chromosomal gene amplifications (copy number alterations), and chromosomal translocations giving rise to gene fusions [
126] (
Table 1).
Table 1. Prevalence and pattern of CMGC kinase subfamily dysregulation in cancer.
|
Kinase Family
|
Deregulation Mechanism
|
Examples of Cancer Types
|
|
CDKs
|
Mutations, amplifications, deletions, and altered expression levels
|
Breast cancer [127,128,129], lung cancer [130], pancreatic cancer [131]
|
|
MAPKs
|
Mutations in pathway components (e.g., RAS, RAF)
|
Melanoma [132,133,134], colorectal cancer [135], lung cancer [48,136]
|
|
DYRK
|
Overexpression
|
Pancreatic [137,138], ovarian cancer [139,140], osteosarcoma [141], rhabdomyosarcoma [142]
|
|
GSKs
|
Altered expression levels, post-translational modifications
|
Glioblastoma [143], pancreatic cancer [144], colon cancer [145]
|
|
CLKs
|
Overexpression, cancer-associated splicing alterations
|
Breast cancer [146], prostate cancer [147], ovarian cancer [148]
|
4. Protein–Protein Interactions of the CMGC Kinases
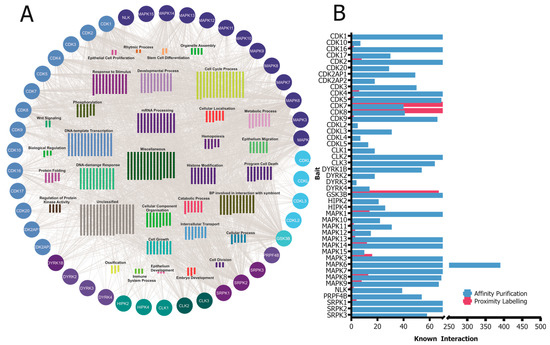
CMGC kinases participate in various protein–protein interactions that are crucial for their activity, functions, and signaling pathways. Understanding these interactions is essential to elucidate the molecular mechanisms governing CMGC-kinase-mediated cellular processes. In this section, we will discuss some of the known interactions involving CMGC kinases and their importance (Figure 2).
Figure 2. (
A) A network of known high-confidence protein–protein interactions of CMGC kinases obtained from the IntAct Molecular Interaction Database are grouped according to their simplified GO process annotation. (
B) Distribution of the number of known interactors of the CMGC kinases based on the affinity purification and proximity labeling detection method. The visualization of the protein–protein interaction (PPI) network was generated by importing the publicly available interaction dataset from
https://www.ebi.ac.uk/intact (accessed on 8 May 2023) protein interaction data into Cytoscape v3.9.
CMGC kinases contribute to the development and progression of various diseases. Therefore, understanding the intricate network of protein–protein interactions involving CMGC kinases can provide valuable insights into the molecular mechanisms underlying their functions and offer potential therapeutic targets for the treatment of diseases associated with their dysregulation. For instance, the development of small molecule inhibitors that specifically target the protein–protein interactions of CMGC kinases could represent a promising approach to modulate their activity and restore normal cellular functions in disease settings [
180].
The development of small molecule inhibitors that target the protein–protein interactions (PPIs) of these kinases has emerged as a promising strategy in cancer therapeutics. Traditional kinase inhibitors target the ATP-binding site of kinases. However, this site’s high conservation across different kinases challenges the development of inhibitors with high specificity. In contrast, the PPI interfaces are often less conserved, presenting an opportunity for creating more specific inhibitors [
183,
184]. Small molecule inhibitors act by obstructing the critical protein interactions necessary for the activation or functioning of these kinases. For instance, inhibitors may preclude a kinase from interacting with its activator or substrate, consequently inhibiting the kinase’s activity and downstream signaling [
185]. In cancer, these inhibitors can restore normal cellular functions by blocking the aberrant signaling engendered by dysregulated kinases. For example, if a kinase is overactive in a cancer cell, promoting uncontrolled cell proliferation, an inhibitor that obstructs this kinase’s interactions could impede or slow down the proliferation [
186]. Considerable progress has been made in the development of such inhibitors. High-throughput screening techniques have enabled researchers to identify small molecules that can bind to the PPI interfaces of CMGC kinases [
185]. Furthermore, advances in structural biology have provided intricate details of these kinases’ 3D structures and their complexes with interacting proteins, aiding in the design of more effective inhibitors [
187]. Nonetheless, challenges persist in developing PPI inhibitors. The interfaces involved in PPIs are often expansive and flat, complicating the ability of small molecules to bind with high affinity. Also, these inhibitors must navigate cellular barriers to reach their intracellular targets [
186].
Most of the studies regarding kinase interactions have focused on a single kinase and its function or dysfunction in a disease. Additionally, the names of the CDK kinases were unified as late as in 2009 [
10], and that might have contributed to the slowness of studies of some these kinases. It is, after all, rather difficult to assign a kinase to this interesting family when the name is very different, such as PFTK1 or CRKRS.
5. Assessment of Kinase Activity
As the function of protein kinases is to transfer a phosphate group, Western blotting with antibodies against a substrate’s phosphoserine, -threonine or -tyrosine is the most common and widely used method to indirectly identify kinase activity. This approach is very labor-intensive, highly dependent on the specificity and sensitivity of the available antibodies, and not directly comparable with high-throughput approaches.
A more sensitive, direct approach to quantitatively measure kinase activity in vitro is to measure the incorporation of radiolabeled phosphate from γ-
33P ATP into the substrate [
205]. A prerequisite to the solution-based in vitro kinase assays is a purified kinase, which in most cases is compared to the kinase dead mutant that abolishes ATP binding. An extensive study of 84 pairs of wild-type kinases and their kinase-dead counterparts [
9] confirmed, using in vitro kinase assays, that indeed the mutated “VAIK” or “HRD” motif is essential for kinase activity. The lysine in the “VAIK” motif is among the most conserved amino acids in all kinases as it is a central catalyzer of the phosphate transfer [
5].
Functional protein microarrays are an efficient ‘omics’-scale tool to detect kinase-substrate interactions [
206,
207]. The commercially available protein microarrays have thousands of purified proteins spotted onto a glass slide, providing an efficient means to identify the phosphorylation of candidate substrates for the kinases of interest when incubated together with radiolabeled ATP [
208]. To date, the only large-scale study utilizing protein microarrays in kinase substrate identification has been performed with yeast protein kinases [
209], although human protein microarrays are nowadays commercially available.