Non-indigenous species are species distributed outside their historic and native range. It has been proposed that a non-indigenous species must go through three stages to become invasive. Firstly, individuals of the species must disperse from their native range to a new area. Secondly, after the introduction, the individuals must survive and reproduce in the new area and become established. Thirdly, once established, the non-indigenous species will increase in number, expand its geographic range, and become a threat to the ecosystem, i.e., become invasive. Thus, the success of invasion depends on the combination of dispersal and demography in a non-native region, and these are affected by the processes of post-dispersal adaptation, genetic diversity, and phenotypic plasticity. However, these processes are not well understood. However, it is well known that invasive species threaten native biodiversity. Based on evidence from metacommunity models, it has been shown that species introductions could disrupt species coexistence, generating extinction debts, especially when combined with other forms of anthropogenic environmental changes. Therefore, the control and eradication of invasive species are essential for the conservation of native species, biodiversity, and ecosystem function, e.g., plant–pollinator networks.
1. Traits Important for Successful Plant Invasion
There has been great interest in determining mechanisms that make the switch from non-indigenous to invasive species happen and in revealing characteristics that contribute to a species’ invasive ability [
9,
10,
11].
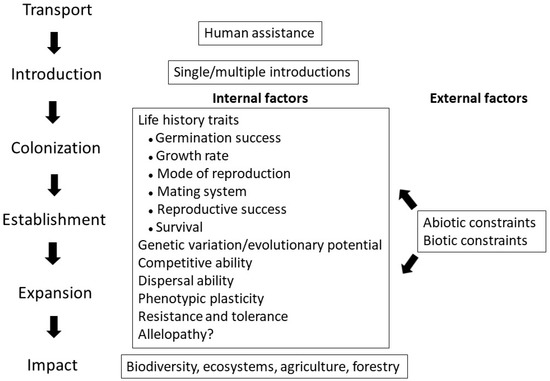
Figure 1 summarizes the process of plant invasion and shows the internal and external factors that affect the process. External factors consist of a range of abiotic and biotic constraints. How well a plant species performs under different conditions depends on a wide range of internal, partly overlapping characteristics. These include multiple life history traits, genetic variation patterns, competitive and dispersal abilities, phenotypic plasticity, resistance, tolerance, and possibly the effect of allelopathic interactions. Control and eradication measures of invasive plants become increasingly difficult as the invasion process progresses. Failure in those actions may result in harmful impacts on the biodiversity and ecosystem functioning, as well as on forestry and agricultural production.
Figure 1. Internal and external factors affect the process of plant invasion from colonization to expansion, which may lead to impacts on biodiversity, ecosystems, forestry, and agricultural production. External factors consist of a range of abiotic and biotic constraints. The role of allelopathy is debated. The figure was prepared by the authors.
One of the questions is whether the success of invaders is driven by post-introduction evolution or high plasticity. The idea that high phenotypic plasticity has contributed to the success of invasive species was proposed in the 1960s [
12]. Indeed, it appears that many invasive plant species are more plastic in several traits than co-occurring non-invasive ones, but this plasticity is only sometimes associated with a fitness benefit based on a large-scale meta-analysis [
10]. However, there are reports providing evidence of the importance of phenotypic plasticity in, e.g., morphological traits [
13], reproductive traits [
14], resource allocation [
15], tolerance against herbivores [
16], light use [
17], and response to altered N supply [
18]. However, investigations on
Plantago lanceolata have indicated that phenotypic plasticity may have initially mediated its adaptation to stressful habitats, while the loss of plasticity associated with secondary invasion may inhibit its further range expansion [
19]. Thus, although there is plenty of evidence for the role of phenotypic plasticity in promoting plant invasion, the evidence is not conclusive.
It has become evident that evolutionary phenomena connected with range expansion drive rapid life-history evolution [
20,
21]. The range expansion itself may be a process that selects for traits that aid invasion success at the range edge, such as enhanced dispersal and colonization, the latter including reproduction success and competitive ability [
11,
22,
23]. In addition, invasive plants with high dispersal capabilities may benefit from increasing habitat fragmentation typically caused by humans. When comparing the effects of different types of life-history traits, it has been suggested that factors associated with evolutionary adaptation and population expansion might determine invasion success and extent, while traits related to the relative competitive ability of invasive species would determine the severity of impacts [
24]. This has implications for the measures used to control invasive plants.
In an investigation on life-history evolution in the invasive honeysuckle
Lonicera japonica, it was discovered that its evolution within the area of invasion has resulted in an enhanced survival and increased growth rate that may drive spread, thus increasing the likelihood of further invasion [
23]. In wind-dispersed plants, a trade-off is expected between dispersal and colonization ability, including germination success and growth that is mediated by seed size; smaller seeds tend to have a better dispersal ability, but possibly a poorer colonization ability [
22]. However, the dispersal ability of the invasive wind-dispersed plant
Gladiolus queinzii Kunze improves towards the range edges in eastern Australia, and this is mediated by a decreased seed size and faster germination time [
11]. Thus, a change towards superior dispersal is not necessarily associated with an inferior colonization ability, even in wind-dispersed plants.
The results on the salt marsh grass
Spartina alterniflora (Loisel.) P.M.Peterson & Saarela support the hypothesis of the evolution of increased competitive ability, in this case through greater aboveground and belowground biomass [
25]. In fact, belowground competition events may turn out to be important for the success of invaders. In experiments in the invasive grass species
Agrostis capillaris L., it was shown that belowground competition can be more important in driving invasive plant impacts than aboveground competition in both low- and high-fertility ecosystems, including those experiencing N enrichment due to global change [
26]. It was found that competition with the invader had large negative impacts on native species’ growth (biomass decreased by half), resource capture (total N content decreased by up to 75%) and even nutrient stoichiometry (tissue C:N ratios of native species increased).
2. Interactions between Genetic Traits and Environmental Conditions Promoting Plant Invasion
The suitability of the environment in the region of introduction is crucial for the establishment and spread of introduced species [
1,
27,
28], as is their genetic composition and adaptability [
4,
29]. Mechanisms involving interactions between genetic traits and environmental conditions that lead to the adaptation of invasive species have been proposed. For instance, inbreeding × environment (I × E) interactions may have a role in invasion success. It has been suggested that a temporary or permanent release from stress in invaded habitats may alleviate the negative effects of insufficient genetic variation on fitness via I × E interactions and that may even generate adaptive genetic changes despite limited genetic variation [
30].
There is evidence that populations spreading through favorable habitats can rapidly evolve high dispersal and reproductive rates, specifically at the expansion front, which further accelerates the spread velocity [
28]. However, spreading populations are likely to encounter stressful conditions somewhere within the expansion range. How evolution during spread in favorable environments affects populations in harsher environments is largely unknown. When comparing evolutionary changes in performance under harsh conditions, i.e., under drought, interspecific competition, and heat stress, in
Arabidopsis thaliana (L.) Heynh. populations, it was discovered that evolution during spread in favorable conditions may either constrain or extend the eventual range limit of species invasion [
28]. A global demographic survey conducted for over 500 plant species showed that populations of invasive plants have superior potential to recover from disturbance compared to non-invasive ones [
31]. Recovery from demographic disturbance was considered to be a measure of transient population amplification, linked to high levels of reproduction.
Global change may reduce evolutionary advantages that native species have developed via adaptation. An investigation into the joint demographic effects of reproductive phenology and warming on the globally invasive thistle species
Carduus nutans L. revealed a substantial shift toward the completion of the life cycle at younger ages [
32]. Demographic modelling projected a 15% increase in this invader’s population growth rate. It was shown that rising temperatures accelerate its population growth by increasing the average size of reproducing individuals, the proportion of individuals that survive to reproduce, and the fraction that reproduce as annuals [
32]. Increased growth and physiological performance under experimental conditions has been observed in the ground-creeping plant
Carpobrotus edulis (L.) N.E. Br, which suggests that climate change would further promote its invasion [
33].
Parthenium hysterophorus L. is a globally invasive weed with significant negative impacts. An analysis on
P. hysterophorus showed that elevated atmospheric carbon dioxide concentration, high ambient temperatures, heatwaves, and droughts will facilitate its establishment and range expansion [
34]. It is evident that climate change could aggravate problems caused by invasive plants and impose increasing management challenges. However, these results are not definite. For instance, a recent meta-analysis found that global change may rather offset than intensify the impacts of biological invasions [
35].
3. Allelopathy and Herbivory Affecting Invasion Success
The allelopathy of plants, i.e., the production of biochemicals that influence the growth, survival, development, and reproduction of other organisms, arises through coevolution [
36]. These compounds may impact plants, animals, and microbes. Plant-interkingdom interactions, especially those with soil-dwelling organisms, have been shown to be important factors in plant health and fitness, possibly providing a competitive advantage [
37]. Allelopathy has been reported to influence the invasiveness of many species, for instance in the common lantana,
Lantana camara L., a species native to tropical America [
38], in the small mimosoid tree,
Leucaena leucocephala (Lam.) de Wit, native to southern Mexico and Central America [
39], and in the Mexican sunflower,
Tithonia diversifolia (Hemsl.) A.Gray, native to Mexico and Central America [
40]. A review of allelopathic information on 524 invasive plant species demonstrated that most of them produce allelochemicals with the potential to negatively affect the performance of native plants [
40]. The released allelochemicals may suppress the regeneration process of other plant species by decreasing their germination and seedling growth and increasing their mortality. Therefore, allelopathy may support the invasive potential of
L. camara,
T. diversifolia, and others, and the formation of dense monospecies stands.
Herbivory can influence the evolution of plants’ chemical traits. Populations that escape selection from their ancestral herbivores may shift resources from defense to competitiveness [
41,
42]. Experiments on the goldenrod
Solidago altissima L. have suggested that a release from herbivory is an important driver of plant adaptation in the invasive range [
43]. However, studies on native and invasive populations of the African daisy,
Senecio pterophorus DC., concluded that herbivory was not a selective factor after invasion [
44]. Instead, the climate was a driver for post-invasion evolution, as suggested by plant adaptations to a drought cline in the native range, the analogous change in plant traits in invaded regions, and the convergence of vegetative traits between non-native and native plants under similar drought conditions.
The Shifting Defense hypothesis gives a somewhat different view to the herbivory question among invasive plants [
43,
45,
46]. Based on this hypothesis, invasive plants are likely to be released from specialist herbivores and, at the same time, they will encounter biotic resistance from local generalist herbivores in their new ranges. This will result in the evolution of a decreased defense against specialist herbivores and an increased defense against generalist herbivores. The meta-analysis conducted by Zhang et al. [
46] indicated that two major mechanisms, i.e., release from specialist enemies and biotic resistance by generalist herbivores, may be pivotal in the evolution of defenses against herbivores. Thus, the analysis provided support for the Shifting Defense hypothesis. Additionally, seed predation can structure plant communities if there is selective foraging. It has been shown, for instance, that rodents prefer to eat the seeds of native grass species instead of the invasive cheatgrass,
Bromus tectorum L. [
47]. Rodent foraging reduced the establishment of each native species by at least 80%, but had no effect on the establishment of cheatgrass.
This entry is adapted from the peer-reviewed paper 10.3390/life13071596