Impaired cognition is the primary symptom of dementia, which can lead to functional disability and reduced quality of life among an increasingly ageing population. Ageing is associated with increased oxidative stress, chronic low-grade systemic inflammation, and endothelial dysfunction, which reduces cerebrovascular function leading to cognitive decline. Chronic low-grade systemic inflammatory conditions, such as obesity, exacerbate this decline beyond normal ageing and predispose individuals to neurodegenerative diseases, such as dementia. Capsaicin, the major pungent molecule of chilli, has recently demonstrated improvements in cognition in animal models via activation of the transient receptor potential vanilloid channel 1 (TRPV1). Capsaicin-induced TRPV1 activation reduces adiposity, chronic low-grade systemic inflammation, and oxidative stress, as well as improves endothelial function, all of which are associated with cerebrovascular function and cognition.
- capsaicin
- cerebrovascular
- cognition
- ageing
1. Capsaicin
| Disease State | Main Findings | Reference |
|---|---|---|
| Cardiovascular | ↓Blood pressure | [97][14] |
| Cancer | Anti-proliferative | [88][3] |
| Neuropathic pain | ↓Painful neuropathy | [98,99][15][16] |
| Adiposity and metabolic derangements | ↑Energy expenditure ↑Fat oxidation ↑Thermogenesis ↑Glucose tolerance ↑Insulin sensitivity ↑Resting metabolic rate ↓Body mass ↓Total cholesterol ↓Triglycerides ↓ Glucose |
[83,100,101,102,103,104][8][17][18][19][20][21] |
Capsaicin: A Brief Overview of Its Role as Anti-Cardiometabolic Disease Treatment
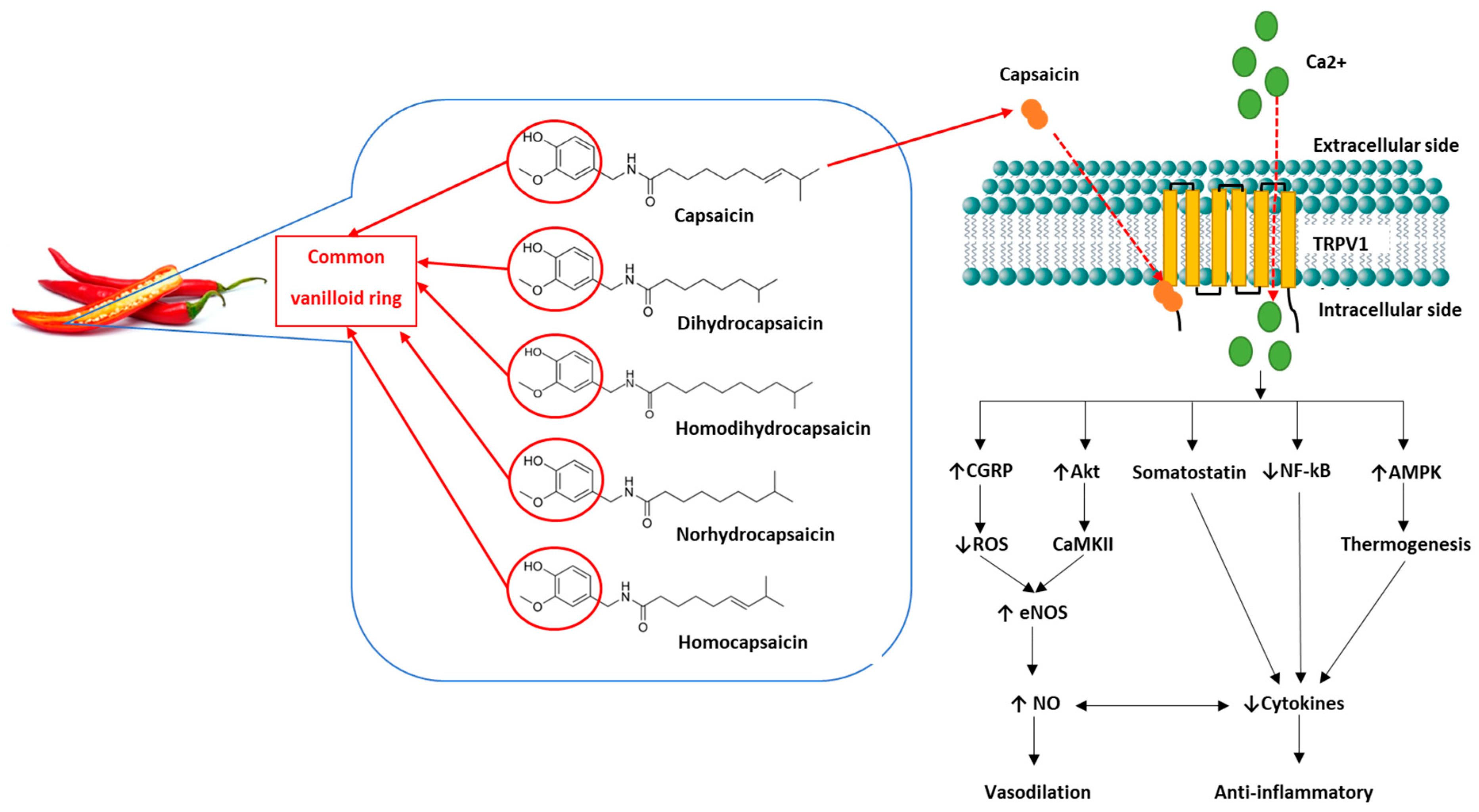
2. The Effects of Capsaicin on Cognition and Cerebrovascular Function
2.1. Cognition in Animal Studies
2.2. Cognition in Human Studies
2.3. Cerebrovascular Function in Animals and Humans
References
- Gaber El-Saber, B.; Alqahtani, A.; Oluwafemi Adeleke, O.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A.; et al. Biological properties, bioactive constituents, and pharmacokinetics of some Capsicum spp. and capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179.
- Rollyson, W.D.; Stover, C.A.; Brown, K.C.; Perry, H.E.; Stevenson, C.D.; McNees, C.A.; Ball, J.G.; Valentovic, M.A.; Dasgupta, P. Bioavailability of capsaicin and its implications for drug delivery. J. Control. Release 2014, 196, 96–105.
- Adetunji, T.L.; Olawale, F.; Olisah, C.; Adetunji, A.E.; Aremu, A.O. Capsaicin: A two-decade systematic review of global research output and recent advances against human cancer. Front. Oncol. 2022, 12, 908487.
- Shrivastava, P.R.; Ravishankar, G.A. Casaicin: A promising multifaceted drug from Capsicum spp. Evid. Based Integr. Med. 2005, 1, 147–166.
- Luo, X.J.; Peng, J.; Li, Y.J. Recent advances in the study on capsaicinoids and capsinoids. Eur. J. Pharmacol. 2011, 650, 1–7.
- Tyagi, S.; Shekhar, N.; Thakur, A.K. Protective role of capsaicin in neurological disorders: An overview. Neurochem. Res. 2022, 47, 1513–1531.
- Tanaka, Y.; Hosokawa, M.; Otsu, K.; Watanabe, T.; Yazawa, S. Assessment of capsiconinoid composition, nonpungent capsaicinoid analogues, in capsicum cultivars. J. Agric. Food Chem. 2009, 57, 5407–5412.
- Panchal, S.K.; Bliss, E.; Brown, L. Capsaicin in metabolic syndrome. Nutrients 2018, 10, 630.
- Yang, D.; Luo, Z.; Ma, S.; Wong, W.T.; Ma, L.; Zhong, J.; He, H.; Zhao, Z.; Cao, T.; Yan, Z.; et al. Activation of TRPV1 by dietary capsaicin improves endothelium-dependent vasorelaxation and prevents hypertension. Cell Metab. 2010, 12, 130–141.
- Ching, L.C.; Kou, Y.R.; Shyue, S.K.; Su, K.H.; Wei, J.; Cheng, L.C.; Yu, Y.B.; Pan, C.C.; Lee, T.S. Molecular mechanisms of activation of endothelial nitric oxide synthase mediated by transient receptor potential vanilloid type 1. Cardiovasc. Res. 2011, 91, 492–501.
- Patowary, P.; Pathak, M.P.; Zaman, K.; Raju, P.S.; Chattopadhyay, P. Research progress of capsaicin responses to various pharmacological challenges. Biomed. Pharmacother. 2017, 96, 1501–1512.
- Yao, X.; Garland, C.J. Recent developments in vascular endothelial cell transient receptor potential channels. Circ. Res. 2005, 97, 853–863.
- McCarty, M.F.; DiNicolantonio, J.J.; O’Keefe, J.H. Capsaicin may have important potential for promoting vascular and metabolic health. Open Heart 2015, 2, e000262.
- Amini, M.R.; Sheikhhossein, F.; Bazshahi, E.; Hajiaqaei, M.; Shafie, A.; Shahinfar, H.; Azizi, N.; Gharehgheshlaghi, H.E.; Naghshi, S.; Fathipour, R.B.; et al. The effects of capsinoids and fermented red pepper paste supplementation on blood pressure: A systematic review and meta-analysis of randomized controlled trials. Clin. Nutr. 2021, 40, 1767–1775.
- Derry, S.; Rice, A.S.; Cole, P.; Tan, T.; Moore, R.A. Topical capsaicin (high concentration) for chronic neuropathic pain in adults. Cochrane Database Syst. Rev. 2017, 1, 1465–1858.
- D’Souza, R.S.; Barman, R.; Joseph, A.; Abd-Elsayed, A. Evidence-based treatment of painful diabetic neuropathy: A systematic review. Curr. Pain Headache Rep. 2022, 26, 583–594.
- Ludy, M.J.; Moore, G.E.; Mattes, R.D. The effects of capsaicin and capsiate on energy balance: Critical review and meta-analyses of studies in humans. Chem. Senses 2012, 37, 103–121.
- Whiting, S.; Derbyshire, E.; Tiwari, B.K. Capsaicinoids and capsinoids. A potential role for weight management? A systematic review of the evidence. Appetite 2012, 59, 341–348.
- Irandoost, P.; Lotfi Yagin, N.; Namazi, N.; Keshtkar, A.; Farsi, F.; Mesri Alamdari, N.; Vafa, M. The effect of Capsaicinoids or Capsinoids in red pepper on thermogenesis in healthy adults: A systematic review and meta-analysis. Phytother. Res. 2021, 35, 1358–1377.
- Jiang, Z.; Qu, H.; Lin, G.; Shi, D.; Chen, K.; Gao, Z. Lipid-lowering efficacy of the capsaicin in patients with metabolic syndrome: A systematic review and meta-analysis of randomized controlled trials. Front. Nutr. 2022, 9, 812294.
- Catalfamo, L.M.; Marrone, G.; Basilicata, M.; Vivarini, I.; Paolino, V.; Della-Morte, D.; De Ponte, F.S.; Di Daniele, F.; Quattrone, D.; De Rinaldis, D.; et al. The utility of Capsicum annuum L. in internal medicine and in dentistry: A comprehensive review. Int. J. Environ. Res. Public Health 2022, 19, 11187.
- Szallasi, A. Capsaicin for weight control: “exercise in a pill” (or just another fad)? Pharmaceuticals 2022, 15, 851.
- Li, B.H.; Yin, Y.W.; Liu, Y.; Pi, Y.; Guo, L.; Cao, X.J.; Gao, C.Y.; Zhang, L.L.; Li, J.C. TRPV1 activation impedes foam cell formation by inducing autophagy in oxLDL-treated vascular smooth muscle cells. Cell Death Dis. 2014, 5, e1182.
- Belza, A.; Jessen, A.B. Bioactive food stimulants of sympathetic activity: Effect on 24-h energy expenditure and fat oxidation. Eur. J. Clin. Nutr. 2005, 59, 733–741.
- Wang, J.; Sun, B.L.; Xiang, Y.; Tian, D.Y.; Zhu, C.; Li, W.W.; Liu, Y.H.; Bu, X.L.; Shen, L.L.; Jin, W.S.; et al. Capsaicin consumption reduces brain amyloid-beta generation and attenuates Alzheimer’s disease-type pathology and cognitive deficits in APP/PS1 mice. Transl. Psychiatry 2020, 10, 230.
- Ouchi, N.; Parker, J.L.; Lugus, J.J.; Walsh, K. Adipokines in inflammation and metabolic disease. Nat. Rev. Immunol. 2011, 11, 85–97.
- Sahin, K.; Orhan, C.; Tuzcu, M.; Sahin, N.; Erten, F.; Juturu, V. Capsaicinoids improve consequences of physical activity. Toxicol. Rep. 2018, 15, 598–607.
- Ma, L.; Zhong, J.; Zhao, Z.; Luo, Z.; Ma, S.; Sun, J.; He, H.; Zhu, T.; Liu, D.; Zhu, Z.; et al. Activation of TRPV1 reduces vascular lipid accumulation and attenuates atherosclerosis. Cardiovasc. Res. 2011, 92, 504–513.
- Wang, Y.; Cui, L.; Xu, H.; Liu, S.; Zhu, F.; Yan, F.; Shen, S.; Zhu, M. TRPV1 agonism inhibits endothelial cell inflammation via activation of eNOS/NO pathway. Atherosclerosis 2017, 260, 13–19.
- Dai, Z.; Li, S.; Meng, Y.; Zhao, Q.; Zhang, Y.; Suonan, Z.; Sun, Y.; Shen, Q.; Liao, X.; Xue, Y. Capsaicin ameliorates high-fat diet-induced atherosclerosis in ApoE(-/-) mice via remodeling gut microbiota. Nutrients 2022, 14, 4334.
- Sun, F.; Xiong, S.; Zhu, Z. Dietary capsaicin protects cardiometabolic organs from dysfunction. Nutrients 2016, 8, 174.
- Wang, J.; Tian, W.; Wang, S.; Wei, W.; Wu, D.; Wang, H.; Wang, L.; Yang, R.; Ji, A.; Li, Y. Anti-inflammatory and retinal protective effects of capsaicin on ischaemia-induced injuries through the release of endogenous somatostatin. Clin. Exp. Pharmacol. Physiol. 2017, 44, 803–814.
- Ebenebe, O.V.; Heather, A.; Erickson, J.R. CaMKII in vascular signalling: “friend or foe”? Heart Lung Circ. 2018, 27, 560–567.
- Kim, S.H.; Hwang, J.T.; Park, H.S.; Kwon, D.Y.; Kim, M.S. Capsaicin stimulates glucose uptake in C2C12 muscle cells via the reactive oxygen species (ROS)/AMPK/p38 MAPK pathway. Biochem. Biophys. Res. Commun. 2013, 439, 66–70.
- Kovacs, G.G. Tauopathies. Handb. Clin. Neurol. 2017, 145, 355–368.
- Mietelska-Porowska, A.; Wasik, U.; Goras, M.; Filipek, A.; Niewiadomska, G. Tau protein modifications and interactions: Their role in function and dysfunction. Int. J. Mol. Sci. 2014, 15, 4671–4713.
- Chen, L.; Huang, Z.; Du, Y.; Fu, M.; Han, H.; Wang, Y.; Dong, Z. Capsaicin attenuates amyloid-beta-Induced synapse loss and cognitive impairments in mice. J. Alzheimers Dis. 2017, 59, 683–694.
- Balleza-Tapia, H.; Crux, S.; Andrade-Talavera, Y.; Dolz-Gaiton, P.; Papadia, D.; Chen, G.; Johansson, J.; Fisahn, A. TrpV1 receptor activation rescues neuronal function and network gamma oscillations from Aβ-induced impairment in mouse hippocampus in vitro. eLife 2018, 7, e37703.
- Du, Y.; Fu, M.; Huang, Z.; Tian, X.; Li, J.; Pang, Y.; Song, W.; Tian Wang, Y.; Dong, Z. TRPV1 activation alleviates cognitive and synaptic plasticity impairments through inhibiting AMPAR endocytosis in APP23/PS45 mouse model of Alzheimer’s disease. Aging Cell 2020, 19, e13113.
- Shiri, M.; Komaki, A.; Oryan, S.; Taheri, M.; Komaki, H.; Etaee, F. Effects of cannabinoid and vanilloid receptor agonists and their interaction on learning and memory in rats. Can. J. Physiol. Pharmacol. 2017, 95, 382–387.
- Pegorini, S.; Braida, D.; Verzoni, C.; Guerini-Rocco, C.; Consalez, G.G.; Croci, L.; Sala, M. Capsaicin exhibits neuroprotective effects in a model of transient global cerebral ischemia in Mongolian gerbils. Br. J. Pharmacol. 2005, 144, 727–735.
- Abdel-Salam, O.M.E.; El-Sayed El-Shamarka, M.; Youness, E.R.; Shaffie, N. Protective effect of hot peppers against amyloid beta peptide and brain injury in AlCl(3)-induced Alzheimer’s disease in rats. Iran. J. Basic Med. Sci. 2023, 26, 335–342.
- Claassen, J.A.; Jansen, R.W. Cholinergically mediated augmentation of cerebral perfusion in Alzheimer’s disease and related cognitive disorders: The cholinergic-vascular hypothesis. J. Gerontol. Med. Sci. 2006, 61, 267–271.
- Herholz, K. Acetylcholine esterase activity in mild cognitive impairment and Alzheimer’s disease. Eur. J. Nucl. Med. Mol. Imaging 2008, 35, S25–S29.
- Chaudhary, S.; Scouten, A.; Schwindt, G.; Janik, R.; Lee, W.; Sled, J.G.; Black, S.E.; Stefanovic, B. Hemodynamic effects of cholinesterase inhibition in mild Alzheimer’s disease. J. Magn. Reson. Imaging 2013, 38, 26–35.
- Nalivaeva, N.N.; Turner, A.J. AChE and the amyloid precursor protein (APP)-cross-talk in Alzheimer’s disease. Chem.-Biol. Interact. 2016, 259, 301–306.
- Rajashri, K.; Mudhol, S.; Serva Peddha, M.; Borse, B.B. Neuroprotective effect of spice oleoresins on memory and cognitive impairment associated with scopolamine-induced Alzheimer’s disease in rats. Am. Chem. Soc. Omega 2020, 5, 30898–30905.
- Viayna, E.; Coquelle, N.; Cieslikiewicz-Bouet, M.; Cisternas, P.; Oliva, C.A.; Sánchez-López, E.; Ettcheto, M.; Bartolini, M.; De Simone, A.; Ricchini, M.; et al. Discovery of a potent dual inhibitor of acetylcholinesterase and butyrylcholinesterase with antioxidant activity that alleviates Alzheimer-like pathology in old APP/PS1 mice. J. Med. Chem. 2021, 64, 812–839.
- Shalaby, M.A.; Nounou, H.A.; Deif, M.M. The potential value of capsaicin in modulating cognitive functions in a rat model of streptozotocin-induced Alzheimer’s disease. Egypt. J. Neurol. Psychiatry Neurosurg. 2019, 55, 48.
- Dworkin, S.; Mantamadiotis, T. Targeting CREB signalling in neurogenesis. Expert Opin. Ther. Targets 2010, 14, 869–879.
- Patanè, S.; Marte, F.; Bella, G.D.; Cerrito, M.; Coglitore, S. Capsaicin, arterial hypertensive crisis and acute myocardial infarction associated with high levels of thyroid stimulating hormone. Int. J. Cardiol. 2009, 134, 130–132.
- Singh, Y.; Gupta, G.; Shrivastava, B.; Dahiya, R.; Tiwari, J.; Ashwathanarayana, M.; Sharma, R.K.; Agrawal, M.; Mishra, A.; Dua, K. Calcitonin gene-related peptide (CGRP): A novel target for Alzheimer’s disease. CNS Neurosci. Ther. 2017, 23, 457–461.
- Jiang, X.; Jia, L.-W.; Li, X.-H.; Cheng, X.-S.; Xie, J.-Z.; Ma, Z.-W.; Xu, W.-J.; Liu, Y.; Yao, Y.; Du, L.-L.; et al. Capsaicin ameliorates stress-induced Alzheimer’s disease-like pathological and cognitive impairments in rats. J. Alzheimers Dis. 2013, 33, 91–105.
- Harada, N.; Narimatsu, N.; Kurihara, H.; Nakagata, N.; Okajima, K. Stimulation of sensory neurons improves cognitive function by promoting the hippocampal production of insulin-like growth factor-I in mice. Transl. Res. 2009, 154, 90–102.
- Bashiri, H.; Hosseini-Chegeni, H.; Alsadat Sharifi, K.; Sahebgharani, M.; Salari, A.A. Activation of TRPV1 receptors affects memory function and hippocampal TRPV1 and CREB mRNA expression in a rat model of biliary cirrhosis. Neurol. Res. 2018, 40, 938–947.
- Avraham, Y.; Grigoriadis, N.C.; Magen, I.; Poutahidis, T.; Vorobiav, L.; Zolotarev, O.; Ilan, Y.; Mechoulam, R.; Berry, E.M. Capsaicin affects brain function in a model of hepatic encephalopathy associated with fulminant hepatic failure in mice. Br. J. Pharmacol. 2009, 158, 896–906.
- Liu, C.-H.; Bu, X.-L.; Wang, J.; Zhang, T.; Xiang, Y.; Shen, L.-L.; Wang, Q.-H.; Deng, B.; Wang, X.; Zhu, C.; et al. The associations between a capsaicin-rich diet and blood amyloid-β levels and cognitive function. J. Alzheimers Dis. 2016, 52, 1081–1088.
- Edvinsson, L.; Jansen, I.; Kingman, T.A.; McCulloch, J. Cerebrovascular responses to capsaicin in vitro and in situ. Br. J. Pharmacol. 1990, 100, 312–318.
- Marics, B.; Peitl, B.; Pazmandi, K.; Bacsi, A.; Nemeth, J.; Oszlacs, O.; Jancso, G.; Dux, M. Diet-induced obesity enhances TRPV1-mediated neurovascular reactions in the dura mater. Headache 2017, 57, 441–454.
- Mishra, G.; Willows, J. Sensory nerve neuropeptide calcitonin gene–related peptide (CGRP) in adipose tissue changes according to metabolic status. Diabetes 2022, 71, 208-LB.
- Xu, X.; Wang, P.; Zhao, Z.; Cao, T.; He, H.; Luo, Z.; Zhong, J.; Gao, F.; Zhu, Z.; Li, L.; et al. Activation of transient receptor potential vanilloid 1 by dietary capsaicin delays the onset of stroke in stroke-prone spontaneously hypertensive rats. Stroke 2011, 42, 3245–3251.
- Marquez-Romero, J.M.; Huerta-Franco, M.R.; Vargas-Luna, M.; Madrigal-Gutierrez, C.A.; Esparza-Hernandez, J.M.; Velazquez-Barcena, M.G. Dose escalation and safety of capsaicin for cerebral perfusion augmentation: A pilot study. Stroke 2021, 52, 2203–2209.
- Laolob, T.; Bunyapraphatsara, N.; Waranuch, N.; Pongcharoen, S.; Punyain, W.; Chancharunee, S.; Sakchaisri, K.; Pratuangdejkul, J.; Chongruchiroj, S.; Kielar, F.; et al. Enhancement of lipolysis in 3T3-L1 adipocytes by nitroarene capsaicinoid analogs. Nat. Prod. Commun. 2021, 16, 1–13.
- Jamornwan, S.; Chokpanuwat, T.; Uppakara, K.; Laorob, T.; Wichai, U.; Ketsawatsomkron, P.; Saengsawang, W. Nitro capsaicin suppressed microglial activation and TNF-alpha-Induced brain microvascular endothelial cell damage. Biomedicines 2022, 10, 2680.
- Wang, C.; Lu, J.; Sha, X.; Qiu, Y.; Chen, H.; Yu, Z. TRPV1 regulates ApoE4-disrupted intracellular lipid homeostasis and decreases synaptic phagocytosis by microglia. Exp. Mol. Med. 2023, 55, 347–363.