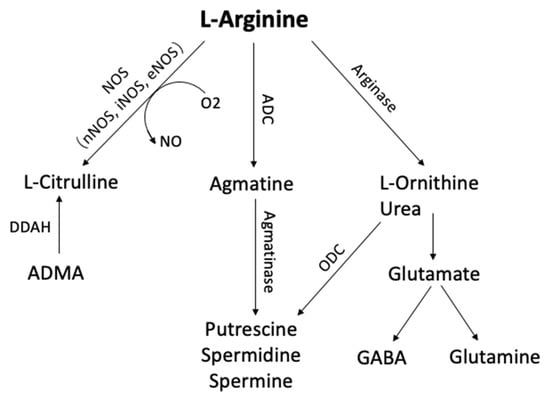
1. Arginine Metabolism
L-arginine is a semi-essential amino acid that can be metabolized to form a number of bioactive molecules
[1][167] (
Figure 1). It is synthesized from proline or glutamate, with the ultimate synthetic step catalyzed by argininosuccinate lyase
[2][168]. L-arginine can be metabolized by arginases, nitric oxide synthases (NOS), and possibly also by arginine decarboxylase (ADC), resulting ultimately in the production of agmatine, ornithine, nitric oxide (NO), or urea
[2][168]. The expression of several of these enzymes can be regulated at transcriptional and translational levels by changes in the concentration of L-arginine itself
[3][169].
Figure 1. Arginine metabolic pathways. L-arginine can be metabolized by phosphatidic acid (PA), nitric oxide synthase (NOS), arginase, and arginine decarboxylase (ADC) to form several bioactive molecules. (ADC, Arginine decarboxylase; ADMA, NG-dimethyl-L-arginine; DDAH, dimethylarginine dimethylaminohydrolase; GABA, γ-aminobutyric acid; ODC, ornithine decarboxylase)
L-ornithine is the arginase-mediated metabolite of L-arginine, with urea as the by-product. L-ornithine can be further metabolized to form putrescine, spermidine, and spermine polyamine, which are essential for normal cell growth and functioning, or via a separate pathway to form glutamine and cell-signaling molecule, GABA
[1][167]. Previous research has reported decreased glutamate and GABA levels in A
lzheimer’s disease (AD
) brains and increased glutamine synthase (GS) levels in the lumbar cerebrospinal fluid of AD patients
[4][5][170,171]. In peripheral organs and also CNS, arginine can also be metabolized by ADC to produce agmatine, a neurotransmitter that plays an important role in the learning and memory process
[6][172].
NO is a gaseous signaling molecule produced by NOS. NO, derived from neuronal NOS (nNOS), plays an important role in synaptic plasticity and learning, and memory
[7][8][9][173,174,175]. Moreover, L-arginine and NO affect the cardiovascular system as endogenous antiatherogenic molecules that protect the endothelium, modulate vasodilatation, and interact with the vascular wall and circulating blood cells
[10][11][12][13][14][176,177,178,179,180].
2. Alanine, Aspartate, and Glutamate Metabolism
Glutamate is the principal excitatory neurotransmitter of the brain
[15][181]. Most neurons and glia are likely to be influenced by glutamate since they have receptors for glutamate. Glutamate is considered the main neurotransmitter of neocortical and hippocampal pyramidal neurons and is involved in higher mental functions such as cognition and memory
[16][182]. Disturbance of excitatory glutamatergic neurotransmission is believed to be associated with many neurological disorders, including AD
[16][182], ischemic brain damage
[17][183], and motor neuron disease
[18][184].
Glutamate receptors can be divided into two classes: ionotropic (N-methyl-D-aspartate, NMDA) and α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)/kainite subtypes and metabotropic
[19][185]. The role of glutamate and glutamate receptors in learning and memory is widely recognized. For instance, NMDA antagonists impair learning and memory while NMDA agonists and facilitators improve memory
[16][182]; likewise, AMPAKines (positive modulators of receptor function) facilitate learning and memory
[20][186]. Circumstantial evidence of the involvement of glutamatergic pathways derives from the well-known role of structures such as the hippocampus in learning and memory
[21][187]. More specifically, lesions of certain glutamatergic pathways impair learning and memory
[22][188]. Moreover, glutamate and glutamate receptors are involved in mechanisms of synaptic plasticity, which are considered to underlie learning and memory
[23][24][25][189,190,191].
3. Purine Metabolism
Purines and pyrimidines are components of many key molecules in living organisms. The primary purines adenine and guanosine and the pyrimidines cytosine, thymidine, and uracyl are the core of DNA, RNA, nucleosides, and nucleotides involved in energy transfer (ATP, GTP)
[26][27][192,193]. Several studies indirectly suggested that purine metabolism has altered in AD. Energy metabolism, which depends on mitochondrial function and ATP production, is markedly altered in AD
[28][29][194,195]. In addition, oxidative damage to DNA and RNA, as revealed by the increase in 8-hydroxyguanosine, is found in the brain samples of AD
[30][31][32][33][196,197,198,199]. Direct alterations of purine metabolism in AD have been detected by metabolomics in postmortem ventricular CSF
[34][200] and in the spinal cord CSF of living individuals
[35][36][37][201,202,203]. Only a limited number of metabolomics studies have been carried out in AD brains
[37][203].
4. Taurine and Hypotaurine Metabolism
Taurine is the second most abundant endogenous amino acid in the central nervous system (CNS) and has multiple roles in our body: thermoregulation
[38][204], stabilization in regulating protein folding
[39][205], anti-inflammatory effects
[40][206], antioxidation
[41][207], osmoregulation
[42][208], and calcium homeostasis
[43][209]. Recently, taurine has shown therapeutic effects as a cognitive enhancer in animal models of non-AD neurological disorders
[44][45][46][47][210,211,212,213]. Taurine protected mice from the memory disruption induced by alcohol, pentobarbital, sodium nitrite, and cycloheximide but had no obvious effect on other behaviors including motor coordination, exploratory activity, and locomotor activity
[44][210]. Intravenously injected taurine significantly improves post-injury functional impairments of traumatic brain injury in rats
[45][211]. The intracerebroventricular (ICV) administration of taurine protects mice from learning impairment induced by hypoxia. Neither beta-alanine nor saccharose was able to mimic the effects of taurine
[46][212]. In streptozotocin-induced sporadic dementia rat models, cognitive impairment and deterioration of neurobehavioral activities are ameliorated by taurine
[47][213].
Taurine also has multiple disease-modifying roles to cease or prevent AD neuropathology. During the development of AD, amyloid-β (Aβ) progressively misfolded into toxic aggregates, which are strongly associated with neuronal loss, synaptic damage, and brain atrophy. An electron microscopy study indicates that taurine slightly decreases β-amyloid peptide aggregation in the brain at a millimolar concentration
[48][214]. Taurine also has anti-inflammatory and antioxidant properties; it can provide protection for neuronal cells and mitochondria from the neurotoxicity of Aβ. By activating GABA and glycine receptors, taurine inhibits excitotoxicity caused by Aβ-induced glutamatergic transmission activation
[49][215].
5. Cholinergic System
As acetylcholine (ACh) plays a vital role in cognitive processes, the cholinergic system is considered an important factor in AD
[50][216]. The brain regions most affected by a loss of elements of the acetylcholine system include the hippocampus, cortex, and entorhinal
[51][217]. Cholinesterase inhibitors are one of the few drug therapies available in the clinic for the treatment of AD, and it was inspired by the fact that cholinesterase inhibitors increase the availability of acetylcholine at brain synapses
[52][218]. The validation of the cholinergic system was seen as an important therapeutic target in the disease.
6. Fatty Acids
Fatty acids are the basic building blocks of more complex lipids and can be classified by the number of double bonds as saturated fatty acids (SFAs) and unsaturated fatty acids. SFAs do not include any double bonds, whereas unsaturated fatty acids contain at least one (monounsaturated fatty acids, MUFAs) or two or more (polyunsaturated fatty acids, PUFAs) double bonds
[53][54][219,220]. Altered unsaturated fatty acids have been associated with AD in multiple studies. The brain is especially enriched with two PUFAs: docosahexaenoic acid (DHA) and arachidonic acid (AA). DHA, as one of omega-3 PUFAs, is the predominant structural fatty acid in the mammalian brain and plays an essential role in brain functioning, especially in cognitive function; DHA levels were lower in AD brains
[55][56][221,222] or plasma
[57][69], and increased intake of DHA from fish or marine oils may lower AD risk
[58][59][60][223,224,225]. AA of the ω-6 fatty acid family appears to play critical mediator roles in amyloid (Aβ)-induced pathogenesis, leading to learning, memory, and behavioral impairments in AD
[61][226]. The levels of free AA have been found to increase in AD patient brain samples
[62][82], whereas the levels of AA in phospholipids are reduced in the hippocampus of AD subjects
[63][227].
7. Glycerolipids
Glycerolipids can be categorized into triacylglycerols (TAG, also known as triglycerides, TG), monoacylglycerol (MAG), and diacylglycerol (DAG) based on the number of acyl groups in the structure. TAG, the most predominant glycerolipids, are esters composed of a glycerol backbone and three fatty acids. TAG levels are found not to be changed in the serum of AD patients when compared to control subjects.
[64][228]. However, MAG and DAG are elevated in both the prefrontal cortex and plasma of AD and MCI subjects in comparison to controls
[65][66][229,230]. Moreover, MAG and DAG are elevated in the grey matter of MCI and AD patients, suggesting that these biochemical changes may play a role in the development of MCI and in the transition from MCI to AD
[67][231].
8. Glycerophospholipids
Glycerophospholipids (GPs), also referred to as phospholipids (PLs), are typically amphipathic and make up the characteristic lipid bilayer structure of biological membranes. Moreover, GPs are the major type of lipids that make up cell membranes and account for 50–60% of the total membrane mass along with cholesterol and glycolipids
[68][232]. GPs include phosphatidylethanolamine (PE), phosphatidic acid (PA), phosphatidylserine (PS), phosphatidylglycerol (PG), phosphatiylcholine (PC), phosphatidylinositol (PI), sphingomyelin (SM), and cardiolipin (CL)
[69][233]. Studies on GP composition indicate that levels of PC, PE, and PI are significantly decreased in neural membranes from different regions of AD patients compared to age-matched control brains
[70][71][72][73][74][75][234,235,236,237,238,239].
Phosphatidylethanolamine (PE) is converted to lysophosphatidylethanolamine (lyso-PE) by phospholipase A2 (PLA2), an important inflammatory mediator that is dysregulated in AD. PLA2 level has been found to be elevated in the human cerebral cortex
[76][240] or decreased in the human parietal and frontal cortex
[77][241]. Moreover, PLA2 influences the processing and secretion of amyloid precursor protein, which gives rise to the β-amyloid peptide, the major component of the amyloid plaque in AD
[77][241]. Moreover, PLA2 has been found to play an important role in memory retrieval
[78][242].
Phosphatidylserine (PS) is the major acidic phospholipid class that accounts for 13–15% of the phospholipids in the human cerebral cortex
[79][243]. PS is known as a “brain nutrient”, as it can not only nourish the brain, but also enhance brain functions such as improving cognition, memory, and reaction force
[80][244]. In six double-blind trials, PS has been found effective for AD. At daily doses of 200–300 mg for up to six months, PS consistently improved clinical global impression and activities of daily living
[81][245]. In milder cases, PS improved orientation, concentration, learning, and memory for names, locations, and recent events. In the largest trial, involving 425 elderly patients (aged between 65 and 93 years) with moderate to severe cognitive decline, PS significantly improved memory, learning motivation, and socialization, suggesting that it has a vital impact on the quality of life of such elderly patients.
Phosphatidylcholine (PC) is an essential component of cell membranes and makes up approximately 95% of the total choline compound pool in most tissues
[82][83][246,247]. Its function is defined primarily by chain length since chain length differences can affect cell membrane fluidity
[84][248]. Three PCs (PC 16:0/20:5, PC 16:0/22:6, and PC 18:0/22:6) have been found significantly diminished in AD patients
[85][249].
Lysophosphatidic acids (LPAs) are phospholipids derivatives that can act as signaling molecules
[86][250]. Ahmad et al.
[87][95] investigated the association between LPAs and CSF biomarkers of AD, Aβ-42, p-tau, and total tau levels overall and with MCI to AD progression. Five LPAs (LPA C16:0, LPA C16:1, LPA C22:4, LPA C22:6, and isomer-LPA C 22:5) correlated significantly and positively with CSF biomarkers of AD, Aβ-42, p-tau, and total tau. Additionally, LPA C16:0 and LPA C16:1 showed associations with MCI to AD dementia progression.
9. Sphingolipids
Sphingolipids, a class of membrane biomolecules, include sphingosine 1-phosphates (S1P), Ceramider (Cers), SMs, and glycosphingolipids, which are vital for maintaining cell integrity and signal transduction processes
[88][251]. Cers, the basic structural units of the sphingolipid class, have been seen as key contributors to the pathology of AD as they are able to affect both Aβ generation and tau phosphorylation
[89][252]. Filippov et al. found elevated levels of ceramides Cer16, Cer18, Cer20, and Cer24 in the brains of AD patients. Two saturated ceramides, Cer (d18:1/18:0) and Cer (d18:1/20:0) were significantly increased in the senile plaques
[90][253]. High ceramide levels were also found in AD serum
[91][254] and CSF samples
[92][255]. The greatest genetic risk factor for late-onset AD is the ε4 allele of apolipoprotein E (ApoE). ApoE regulates the secretion of the potent neuroprotective signaling lipid S1P
[93][256]. S1P is derived by phosphorylation of sphingosine, catalyzed by sphingosine kinases 1 and 2 (SphK1 and 2). SphK1 positively regulates glutamate secretion and synaptic strength in hippocampal neurons. Reduced levels of S1P have been found in AD brains compared to controls
[93][94][256,257]. All these studies mentioned above suggested that sphingolipid metabolism plays a critical role in AD pathology.
10. Cholesterol and Cholesteryl Esters
Despite the brain occupying only 2% of total body weight, it contains 25% of the body’s cholesterol. Due to the BBB, cholesterol metabolism in the CNS is largely separated from that in the periphery and cholesterol is de novo synthesized in the CNS
[95][258]. Studies have found that brain cholesterol was significantly increased in AD patients than in controls
[96][97][259,260]. Moreover, cholesterol showed abnormal accumulation in the senile plaques of the human brain, a hallmark neuropathological feature of AD
[98][261].