Unlike in animals, dioecy among flowering plants is a rare phenomenon. The vast majority of angiosperm species have a bisexual flower that combines male (androecium) and female (gynoecium) reproductive organs. However, about a quarter of species have dioecious flowers, which can be located within the same plant (monoecious) or on different plants (dioecious). TIn recent years, the development of sequencing techniques, bioinformatics, and molecular biology has led to an increase in interest in the sex determination mechanisms among plants. It is noteworthy that a significant number of dioecious plants have economic value. At the same time, dioeciousness often complicates the growing process. This fact increases the relevance of studies on dioecious crops. In this review, we attempt to summarize the current information on sex chromosomes and the mechanisms of sex determination in dioecious plants, concentrating on species with agricultural importance.
- dioeciousness
- flowering plants
- monoecious plant
- dioecious plant
- sex chromosomes
1. Introduction
2. Features of the Dioecious Crop Cultivation
2.1. Sex Chromosomes and Sex Determination in Herbaceous Crops
2.1.1. Asparagus
This is one of the first dioecious plants in which sex determination was studied [9][47]. Asparagus plants are diploid, 2n = 2x = 20, and their sex chromosomes are homomorphic and do not visually differ from each other [10][11][48,49]. The female plants are homogametic and have XX chromosomes (also known as genotype mm) in their karyotype, while the male plants are heterogametic (XY\Mm karyotype). At the same time, the homogametic karyotype YY (MM), the so-called supermale, is viable, has fertile pollen, and is androdiecious in phenotype—that is, it has male and hermaphrodite flowers. The viability of the YY homozygote indicates the early stages of the dioecy evolution path with the young Y chromosome [12][50]. The mechanism of sex regulation in asparagus is one of the most studied among dioecious plants. A small M locus (835 kb in size) located on homomorphic sex chromosomes is responsible for sex determination in this species [13][12]. The development of male and female reproductive organs requires the presence of two key genes located on the sex chromosomes. These are the gene that suppresses female function (SOFF) and the gene that promotes the development and functioning of the tapetum (aspTDF1). Transcriptome analysis revealed more than 500 differently expressed genes in male and female plants [14][51]. Asparagus DNA methylation was studied using whole-genome BS sequencing (bisulfite sequencing) [15][52]. A difference was found in the DNA methylation containe d in male and female flowers. However, no difference in methylation was found in the genes responsible for the formation of flowers.2.1.2. Sorrel
The Rumex genus is one of the model groups for studying sex determination in plants [16][72]. Therefore, sex determination and sex chromosomes are widely studied in a number of Rumex species. Among these species, there are various systems for determining sex and sex chromosomes (completely hermaphroditic species, monoecious species with XY chromosomes and an active Y chromosome, and species with X:A balance sex determination, including multichromosomal species with the XY1Y2 system). Species such as R. acetosa, R. acetosella, R. hastatalus, and R. suffruticosus are dioecious and widely studied. The R. acetosa karyotype consists of 12 autosomes and sex chromosomes (XX in female plants and the XY1Y2 multichromosomal system in male plants) [17][18][19][20][73,74,75,76]. The sex chromosomes are the largest in the karyotype. Two Y chromosomes contain 26% of the total DNA. Sorrel Y chromosomes are heterochromatic and rich in repetitive DNA sequences [21][77]. In 1999, a group of tandem repeats specific to male sex chromosomes were isolated and called RAYSI (Rumex acetosa Y-chromosome-specific I) [22][78]. After that, a number of satellite DNA repeats specific for sorrel Y chromosomes were described. The repeat sequences of RAYSII and RAYSIII are AT-rich and show about 60% homology with RAYSI [17][73]. Repeat satDNA RAE180 is localized on two Y chromosomes and is also present on a pair of autosomes. [23][79].
2.1.3. Spinacia
Currently, the spinach genome has been sequenced, including by PacBio and Hi-C scaffolding. The sex determination region (SDR) was determined; it is located on chromosome 1 or 4 (according to other sources) [24][28]. It has been shown that the size of the SDR is 24.1 Mb on the Y chromosome and 13 Mb on the X chromosome. At the same time, the SDR on the Y chromosome contains inversions (14.1 Mb) and duplications (10 Mb) that are specific for male plants [25][89]. Studies have shown differences in morphological traits between male, female, and monoecious spinach plants. Vitale et al. (1985) showed that male and female plants significantly differ in height and the biomass of roots and the vegetative part while monoecious (cosexual) plants differ in the studied traits between both male and female plants [26][90]. Another study revealed differences in the response of spinach plants of different sexes (for example, in yield) when fertilized with selenium. Female plants were more productive and contained fewer heavy metals than male plants when selenate was applied. In addition, the contents of vitamin C, chlorophyll and carotenes increased in female plants [27][91].2.1.4. Hemp
Hemp is one of the few annual agricultural herbaceous plants that are dioecious in nature. In addition, male and female cannabis plants differ in habit. The dioecy of hemp seriously affects its agricultural practices. Male and female plants differ in ripening time. Female plants are leafier, have more lignified cell walls due to the better (compared with male plants) accumulation of lignin, bloom later, and stay green for longer [28][102]. These features lead to different levels of fiber quality when growing hemp as a textile crop. Cultivation technology requires expensive and time-consuming manual weeding. When growing hemp for oil, more female plants and fewer male pollinating plants are required. When growing female cannabis plants for medicinal purposes, pollination should not be allowed, since unpollinated flowers are characterized by a higher THC content. In this case, the presence of male plants in the population is undesirable. In this regard, the cultivation of monoecious varieties is more cost-effective [29][103]. The nature of hemp’s monoecy is not yet fully understood. A number of studies have shown that monoecious varieties are female in their genetic nature and have an XX karyotype [30][104] and a genome size similar to female plants [31][105]. Male plants have 18 autosomes and XY chromosomes in their karyotype. At the same time, sex is determined according to the balance mechanism. To recognize the sex of plants at any stage of cultivation, molecular genetic markers have been developed in hemp [32][33][34][29,106,107]. However, due to the annual nature of the culture, it seems more promising to create monoecious varieties. However, the ratio of male to female flowers in monoecious plants, as well as the sex of cannabis plants in general, is a very unstable trait. There have been cases of the appearance of flowers of the opposite sex in dioecious plants, as well as sex changes under the influence of environmental factors [35][36][37][38][108,109,110,111]. In this regard, a detailed understanding of the mechanisms of the occurrence of dioecy in hemp, as well as the potential opportunity to influence this trait, is of undoubted agronomic importance.2.1.5. Betel
Betel rarely blooms, but there have been reports of economically significant differences in the varieties of male and female plants [39][30]. There is very little information about the chromosome numbers and the level of ploidy of this species. The main number of chromosomes of this species is x = 13; however, polyploid rows are reported in this culture. Apparently, plants with a high level of ploidy are mainly grown. It was shown that female plants are tetraploid and have 52 (2n = 4x = 52) chromosomes in their karyotype, while male plants are triploid and have 39 chromosomes (2n = 3x = 39) [40][119]. Sex markers have been created for betel plants, mainly to accompany the breeding process, which is currently not very developed due to the complexity in the biology of the culture [39][41][42][30,120,121].
2.1.6. Yams
Yams (Dioscorea rotundata) are one of the most important crops in a number of countries, primarily in the African continent, where Nigeria, Ghana, Côte d’Ivoire, and Benin are considered leaders in the production of yams [43][46]. The plant is grown for the production of tubers, which are then used for food. Normally, D. rotundata is a dioecious species; however, cases of monoecious occurrence have been observed. Despite the huge role of this crop in African countries, the selection of this species is difficult, largely due to its dioecious nature and problems with flowering (it is predominantly a vegetatively propagated crop) [44][45][122,123]. In addition, a change in the sex of plants under the influence of the environment is repeatedly observed during the growing season, which also complicates the breeding process of this crop [46][124]. The identification of sex chromosomes in yams is also difficult [47][125]. However, using modern sequencing techniques, the ZZ/ZW sex determination system has been demonstrated [47][125]. In 2020, the first yam genome obtained as long reads using the Oxford Nanopore Technologies technique was presented [48][126].
2.2. Sex Chromosomes and Sex Determination of Perennial Tree Crops Grown for Fruits
Most dioecious species are perennial tree crops [49][129]. One of the differences between perennial tree crops and annual and perennial herbaceous crops is a long juvenile period—the reproductive stage often only occurs by the age of 5–8. This feature causes difficulties in growing such plants, since most perennial tree crops are cultivated for the fruits and seeds that appear from the ovary located on the female plants. Revealing the mechanisms of sex determination and the early diagnosis of the sex of such crops are critical tasks for production and breeding. The accumulation of repetitive DNA sequences in sex chromosomes may contribute to the development of sex-associated molecular markers, which is especially important for agricultural plants with a long juvenile period. In cultures such as papaya (Carica papaya L.), lychee (Litchi chinensis Sonn.), rambutan (Nephelium lappaceum L.), and longan (Euphoria longan Steud.), the sex determination system is complex. There are not only dioecious plants but also hermaphroditic, gynodioecious, and monoecious plants in the population.2.2.1. Papaya
The papaya sex chromosomes were the second to be fully sequenced after the human sex chromosomes [50][137]. They are evolutionarily young and outwardly practically homomorphic [51][52][11,138]. One of the features of papaya is the presence of hermaphroditic plants—in fact, it is a trioecious species, not a dioecious species. There are two types of Y chromosomes in this species. The first type is represented by the Y chromosome, which is found in male plants. The second type is represented by the Yh chromosome, which causes the hermaphroditism of flowers. At the same time, the YY, YYh, and YhYh genotypes are lethal [53][54][33,139]. The non-recombining region on the Y chromosome of papaya was formed as a result of two inversions of retrotransposons and is 8.1 Mb, while the size of the analogous region of the X chromosome is 3.5 Mb. All genes in non-recombining regions have been annotated [55][140]. Wild papaya is generally dioecious but cultivated as a rule of the gynodioecy cultivar [56][131]. It has been shown that evolutionarily hermaphroditic plants have been fixed for about 4000 years during domestication [57][141]. In addition to differences in the ease of cultivation (when growing hermaphroditic plants, male pollinating plants that do not produce fruits are not needed) between female and hermaphroditic plants, there is dimorphism in the shape of the fruits. Fruits from hermaphrodite flowers are more elongated and commercial producers prefer them, rejecting the more rounded fruits of female plants [50][137]. However, plants can change the sex of their flowers during the growing process. Hermaphroditic flowers can stop the development of carpels, turning into functionally male flowers under the influence of environmental factors such as drought or high temperatures [58][59][60][142,143,144].
2.2.2. Vitis
It has been shown that the emergence of hermaphroditic forms occurred about 6000–8000 years ago at the time of the domestication of wild grapes [5][61][62][5,148,149]. Like papaya, grapes have three sex-determining loci—Y (M) is male, X (F) is female, and Yh (H) is hermaphroditic. Unlike papaya, the HH, HM, or MM genotypes are viable in grapes. Three sex-linked grape genes (ViviPLATZ, VviFSEX, and APRT3) have been described, and the expression of each differs in hermaphroditic plants [62][149].
2.2.3. Diospyros
It is an autopolyploid species with 2n = 90 or 135 chromosomes (2n = 6x; 2n = 9x; x = 15) [63][152]. Despite the presence of at least one Y chromosome in the karyotype, this species is usually monoecious or has completely female plants, with rare male flowers [64][34]. Recent studies have shown that in the dioecious diploid species D. lotius L. (a close relative of D. kaki), the OGI gene located on the Y chromosome is responsible for sex formation. It encodes a small RNA that targets the autosomal MeGl gene, regulating anther fertility in a dose-dependent manner. In polyploid species (D. kaki, in particular), there is an insertion of a retroelement, named Kali, in the promoter region of the OGI gene that prevents the synthesis of small RNA and promotes the development of monoecy.
2.2.4. Pistachio
Pistachio (Pistacia vera L.) is an economically significant crop for Turkey, Iran, the USA, and a number of other countries [43][46]. It is a dioecious plant species, where females are needed in production but male plants are needed for pollination. There are also monoecious cultivars [65][66][153,154]. From the point of view of the evolution of sex and sex chromosomes, this crop is interesting regarding the ZZ/ZW (2n = 30) system of sex chromosomes [67][155]—that is, the heterogametic sex of the pistachio is female, like birds or some insect species. This system of sex chromosomes is rare in plants. A recent study on sequencing and assembling the genomes of the male and female plants showed that the pistachio W sex chromosome arose from three successive inversions [68][156].2.2.5. Hippophae
he sea buckthorn karyotype consists of 24 chromosomes (22 autosomes and a pair of X/Y sex chromosomes) [69][163]. In the genome (about 2.6 pg in size), researchers have observed a uniquely large number of satellites that may turn out to be good cytogenetic markers [69][70][71][163,164,165]. The development of molecular markers to determine the sex of sea buckthorn in the early stages of cultivation has been quite successful, but the experiments have shown the unstable operation of these markers, which may be due to sea buckthorn’s polymorphism [72][73][74][166,167,168]. Morphologically, male and female plants can only be distinguished by their generative buds after flowering age has been reached. At the same time, mainly female plants are needed in agricultural production. Additionally, breeding is carried out independently for male pollinating plants and female plants [75][169]. These facts demonstrate the need for a deeper study of the genetic determination of the sex in sea buckthorn.
2.2.6. Humulus
Humulus lupulus L. is a dioecious woody vine of the Cannabaceae family that is of great importance in brewing. Hops are grown for the female buds, which contain substances that create the characteristic taste, aroma and bitterness of beer. Due to the popularity of this drink, hops are cultivated on every continent except Antarctica. The hop karyotype consists of 18 autosomes and a pair of sex chromosomes (XX/XY). The Y chromosome is the smallest in the karyotype, which may indicate its degeneration and evolutionary antiquity [76][77][78][172,173,174]. In commercial cultivation, the female varieties are vegetatively propagated. A very important task is to prevent the pollination of inflorescences, since setting seeds spoils the taste of beer [79][175]. Hop is a wind-pollinated crop with light pollen, so just one male plant in the area can cause a lot of economic damage [80][176]. To prevent pollination, all male plants in a nursery and nearby wild populations must be eradicated. As a part of this procedure, molecular genetic markers are used to identify sex in the early stages of ontogenesis [81][82][83][38,177,178]. In hop production, seedless triploid varieties are also cultivated. They are more productive and have a high growth rate [84][85][179,180].
2.2.7. Date Palm
The date palm (2n = 2x = 32) is one of the oldest dioecious crops. As with other crops grown for fruit, its production primarily requires female date palm plants. Males are required in smaller numbers for pollination (a ratio of 1:20 is used, i.e., one male plant pollinates 20 females). At the same time, the date palm blooms in the 5–6th year of cultivation—that is, it is not possible to morphologically detect the sex of a tree before this age. Many molecular markers have been developed to identify the sex of the date palm in the early stages of ontogenesis. The genetic mechanisms of sex determination in the date palm have been recently identified [86][87][88][89][90][181,182,183,184,185]. An association of two genes, presumably associated with sex, was found on chromosome 12 [91][186].2.2.8. Myristica
Nutmeg (Myristica fragrans Houtt., 2n = 38 of 44, according to different communications [92][191]) is a tropical evergreen tree grown in Asia and Africa for its fruits. It is widely used around the world as a spice [93][192]. The world production of nutmeg is estimated at 139,000 tons [43][46]. In agricultural production, female plants are needed more. Attempts to create molecular markers have been made to identify sex in the early stages of vegetation [94][95][96][40,193,194]. However, sex chromosomes in M. fragrans have not been found.2.2.9. Actinidia
Morphologically, the flowers look bisexual, but female plants form flowers with sterile pollen while male plants have an underdeveloped and non-functioning pistil [97][197]. However, some species sometimes have bisexual flowers. Hermaphroditic flowers have been observed in A. arguta, A. chinensis, A. deliciosa, and A. eriantha Benth. [98][196]. Kiwifruit has only been cultivated for just over 100 years, but it has achieved great popularity around the world, and its production is about 4 million tons per year [43][46]. Polyploidy is widespread in species of the Actinidia genus [99][198]. Chromosomal numbers range from 2n = 58 to 2n = 174 (x = 29) [100][199]. Polyploidy does not appear to affect sex in kiwifruit. Male plants have at least one Y chromosome in the karyotype, and female plants have XX [101][102][200,201]. The sex chromosomes are small and homomorphic, with a small SDR [103][104][41,202]. The mechanism of sex determination has been widely studied. This mechanism is based on two genes (the Shy Girl gene (SyGI), the dominant suppressor of carpel development, and the Friendly Boy gene (FrBy), expressed in tapetum cells) [105][15]. Currently, the kiwifruit genome is actively being studied, and genetic maps and molecular markers are being created to aid the breeding of this crop [106][107][108][109][203,204,205,206]. However, hermaphroditism can be determined by additional genetic factors, which creates difficulties for sex determination based on molecular markers [109][206].
2.2.10. Ilex
In this species, sexual dimorphism has been repeatedly shown in the intensity of photosynthesis, leaf area, and higher mortality of female plants. Thus, male plants have a larger leaf area than female plants, and the intensity of photosynthesis is higher in female plants, while the leaves of male plants make the taste of the drink less bitter [110][111][112][113][130,212,213,214]. Like Actinidia, I. paraguariensis is functionally dioecious, with morphologically hermaphroditic flowers, and one of the sexes is sterile or abortive [114][215]. From a genetic point of view, this species is poorly studied. Heteromorphic sex chromosomes have not been identified. Molecular markers for marker-associated selection and sex detection have been developed [115][116][117][216,217,218].2.2.11. Chinese Bayberry
Chinese bayberry (Morella rubra Lour.) is widely cultivated in China for fruits. It is the only edible species of the Myricaceae family [118][219]. The chromosome number is 2n = 16, and the heterogametic sex is presumably female (ZW chromosome system) [119][220]. A comparison of the sequenced genomes of male and female plants revealed a small region (59 kb) on chromosome 8 that is specific to female plants [120][42]. The differential expression of some genes, presumably associated with the development of sex in this species, was also shown [121][221].
3. Conclusions
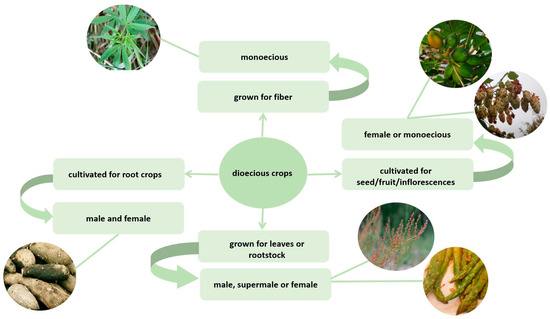
According to Figure 1, dioeciousness has a greater or lesser effect on crop cultivation. It is most noticeable on fruit trees. Gardeners strive to grow as many female plants as possible due to the fact that females produce the target product. However, male plants are necessary for pollination and cannot be completely abandoned. However, the maximum effective sex ratio differs depending on the plant and the type of pollination (insect pollination or wind pollination). For example, sea buckthorn shows the best yield in a planting pattern where one mixed row of male and female plants falls between two rows of female plants (in a mixed row, male plants comprise one in five plants) [122][160]. The role of the pollinator in the formation of economically valuable traits cannot be directly assessed. In addition to dioecy, a long juvenile period also affects the breeding of tree crops. Half of the population during seed propagation comprises male plants. This fact leads to the loss of labor, time, and other resources that are spent on growing males before flowering. Therefore, sex detection using molecular markers is often used in the cultivation of perennial fruit crops. Additionally, the use of monoecious and parthenocarpic forms may be relevant in such cases. At the same time, male plants are only used in the breeding of some other plants, such as hops. The pollination of industrial plantings is unacceptable. The spatial isolation of hop plantings and the early complete culling of male plants is necessary. In plants in which the vegetative part is used, the role of dioecy is not so obvious and it seems that plants of any sex can be used. However, the different maturation times of male and female plants, as in hemp, or the different yields of male and female plants, as in asparagus, can shift the interest of producers to monoecious forms or plants of a specific sex. Thus, researchers should consider the final purpose of the cultivated crop, as well as its biological characteristics in general, not only focusing on the genes for the formation of pistils or stamens. Humans have a significant impact on the entire environment, and dioecious cultures are no exception.