Biofilms enable pathogenic bacteria to survive in unfavorable environments. As biofilm-forming pathogens can cause rapid food spoilage and recurrent infections in humans, especially their presence in the food industry is problematic. Using chemical disinfectants in the food industry to prevent biofilm formation raises serious health concerns. Further, the ability of biofilm-forming bacterial pathogens to tolerate disinfection procedures questions the traditional treatment methods. Thus, there is a dire need for alternative treatment options targeting bacterial pathogens, especially biofilms. As clean-label products without carcinogenic and hazardous potential, natural compounds with growth and biofilm-inhibiting and biofilm-eradicating potentials have gained popularity as natural preservatives in the food industry. However, the use of these natural preservatives in the food industry is restricted by their poor availability, stability during food processing and storage. Also there is a lack of standardization, and unattractive organoleptic qualities. Nanotechnology is one way to get around these limitations and as well as the use of underutilized bioactives. The use of nanotechnology has several advantages including traversing the biofilm matrix, targeted drug delivery, controlled release, and enhanced bioavailability, bioactivity, and stability. The nanoparticles used in fabricating or encapsulating natural products are considered as an appealing antibiofilm strategy since the nanoparticles enhance the activity of the natural products against biofilms of foodborne bacterial pathogens.
- antibiofilm
- biofabrication
- foodborne pathogens
- nanocarrier
- natural preservatives
1. Introduction
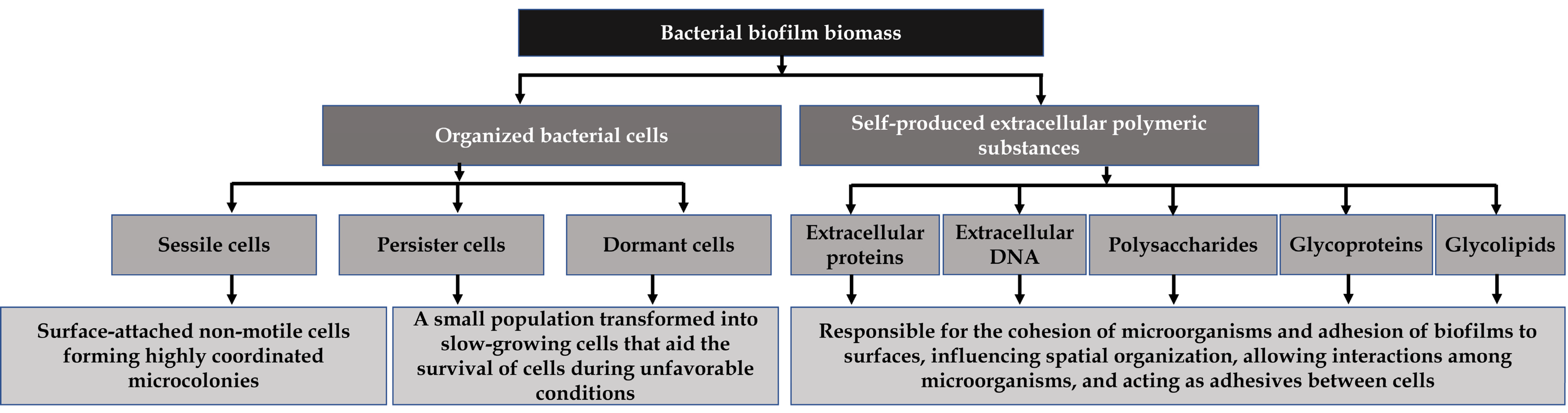
2. Significance of Pathogenic Bacteria and Their Biofilms in the Food Industry
According to earlier reports, materials typically found in the food industry, such as Buna-N, Teflon seals, rubber seals, stainless steel, aluminum, and glasses, as well as food-processing units like heating exchangers, cooling towers, conveyor belts, wastewater pipes, etc., can harbor pathogenic microbes like Escherichia coli, Staphylococcus aureus, Listeria monocytogenes, Campylobacter jejuni, Salmonella spp., Bacillus cereus, and Pseudomonas spp. in the form of biofilms [6,14][6][14]. In food-processing facilities, biofilms have been a critical cause of food deterioration related to economic losses and food safety problems, leading to several outbreaks [6,26][6][15]. Typically, microbes related to the food industry such as B. cereus, Enterohemorrhagic E. coli, L. monocytogenes, and S. enterica, are predominantly present in the mixed biofilm state [7]. Moreover, B. cereus forms endospores that can withstand pasteurization conditions and make robust biofilms [29][16]. The enterotoxin (hemolysin) of B. cereus causes various illnesses, namely diarrhea and abdominal pain [7]. This kind of toxin is stably present in foods with higher butterfat content and in High-Temperature-Short-Time pasteurized food products [30][17]. In the case of enterohemorrhagic E. coli, transmission occurs via ingesting contaminated fruits, vegetables, meat, milk, and distilled water. The E. coli strain O157:H7 forms thick biofilms on stainless steel and borosilicate glass but not on polypropylene surfaces [31][18]. It can potentially secrete Shiga toxins (STEC), spawn enterohaemorrhagic gastroenteritis, and cause watery diarrhea and blood in feces [32][19]. Compared to other foodborne pathogens, L. monocytogenes is a deadly pathogen that accounts for the increased mortality rate [33][20].3. Natural Products as a Promising Alternate
The emergence of drug-resistant microbes is mushrooming at an alarming rate. Compared with other approaches, using natural bioactives could be a promising alternative that inhibits pathogenic bacterial growth and biofilm formation. Consequently, introducing synthetic or unknown ingredients in a food product to hamper microbial incidences causes consumers to consider the product harmful to their health or body. Therefore, various food additives, including genetically engineered products, artificial sweeteners, taste enhancers, food colorants, and preservatives, contribute to the negative reputation of the product. As a result of this tendency, the food industry is considering natural alternatives to synthetic ingredients.3.1. Natural Bioactives as Preservatives
Several natural sources like plants [13], animals [36][21], microorganisms [37][22], bacteriocins [38][23], and even bacteriophages [39][24] are being exploited for the identification of bioactive leads against the growth and biofilm formation of foodborne pathogens, which consequently ended up with an overabundance of compounds [37,40][22][25]. Phytocompounds are one of the bioactive secondary metabolites that might be used as natural food preservatives. Herbal and plant medicines continue to garner interest as potential therapeutics.
Essential oils are highly concentrated, volatile, hydrophobic chemicals present in a wide variety of plants. The hydroxyl groups of essential oil components, such as those in thymol, carvacrol, and eugenol, react with the phospholipid bilayer of microorganisms resulting in leakage of ions, nucleic acids, and ATP and water imbalance, leading to cell death [40][25]. The said compounds at sub-MIC were reported to target the biofilm formation of bacterial pathogens in a concentration-dependent manner.
Furthermore, the bioactives from thyme and rosemary plants reduce the biofilm growth of L. monocytogenes and garlic extract can prevent quorum sensing (QS) signaling in multidrug-resistant bacterial pathogens [40,51][25][26]. In addition, flavones under the flavonoid class form a complex with the components of the bacterial cell wall and impede cell adherence and proliferation. In this regard, the genes Staphylococcus accessory regulator (sarA) and intercellular adhesins (ica) are both downregulated by baicalein to suppress the virulence regulation of S. aureus [52][27].
Glycolipid is a biosurfactant with potential anticancer and antibacterial effects and currently has a wide variety of therapeutic uses [57][28] including in the pharmaceutical, food, and petroleum sectors. Sophorolipid is one of the glycolipids produced by the yeast Starmerella bombicola that has antibacterial and antibiofilm properties against foodborne pathogens such as C. jejuni, E. coli, Listeria spp., and Salmonella spp. [58,59,60][29][30][31].
3.2. Challenges in the Use of Natural Preservatives for Preservation
Natural bioactives possess tremendous potential against biofilm-forming bacterial pathogens. However, a plethora of challenges are associated with the usage of natural bioactives as preservatives. Gram-negative bacteria are rich in lipopolysaccharide (LPS), making it tricky for phenolic compounds to penetrate the cell wall [99][32]. Hence, the catechin monomers from grape seed extract showed increased antibacterial activity towards Gram-positive than Gram-negative bacteria [100][33]. Similarly, bacteriocins cannot penetrate the LPS of Gram-negative bacteria [101][34]. The first obstacle to overcome is the extraction of bioactive compounds from natural sources. The extraction technique must be chosen carefully to preserve the quantitative and qualitative properties of the bioactive compounds [102][35]. Since natural food preservatives need extraction and additional refining procedures, they often increase production costs. Thus, natural food preservatives became more costly than their synthetic counterparts [103][36].4. Nanotechnology Approach
Nanotechnology is a promising technology that has the ability to convert an individual particle to one billionth of its original size. The converted particles are nano-sized (1–100 nm), have a large surface area and mass ratio, and are highly reactive, making them completely different from the exact composition of the bulk material [107][37]. The converted nanoparticles have many advantages, including an increased impact against the target pathogenic microorganisms with multiple functional sites. The exact antibacterial mechanism of nanoparticles has not been entirely elucidated. Many studies have suggested possible mechanisms of action. The absorption of nanoparticles into the cell membrane and the subsequent disintegration are the initial steps involved in the antibacterial mechanism of nanoparticles [108][38]. Following absorption and disintegration, the cell-penetrating nanoparticles target bacterial growth through intracellular content leakage, generation of reactive oxygen species, impairment of the electron transport system, inactivation of efflux pumps, and most importantly, interference with the enzymatic and metabolic activities of the cell [108][38].
4.1. Techniques for Targeting Biofilms through Natural Product Delivery
4.1.1. Biofabrication of Nanoparticles with Antibiofilm Activity Using Surface Functionalization
The surface functionalization of the nanoparticles with phytochemicals or other natural products enhances the bioactivity of the functionalized product. The covalent or non-covalent attachment of the natural product on the surface of the nanoparticles can enhance their solubility, and thereby, its antibiofilm potential. Moreover, the nanoparticle’s penetration enhances the local concentration of the compound inside the biofilm [116][39]. In this regard, metallic nanoparticles have been vastly exploited. Metallic nanoparticles such as gold, silver, copper, and others, synthesized via biological methods (i.e., through plant extracts and natural compounds) frequently have bioactives or compounds in the plant extract as a capping agent [117][40]. Similar to metallic nanoparticles showing intrinsic activity, these nanoparticles act as a carrier for the bioactives that are physically surface-functionalized. The syntheses of nanoparticles often utilize a salt that is exposed to a redox reaction caused by natural extracts or bioactive compounds under specified pH conditions. The same natural substance is responsible for maintaining the freshly prepared nanoparticle’s surface potential and rendering them with antibacterial/antibiofilm activity after the reduction procedure. As a consequence, the oxidized derivative of the active component caps the nanoparticles [118][41]. Nanoparticles generally have desirable qualities, such as the ability to change surface characteristics and stability. Meanwhile, selecting phytocompounds and natural products is crucial in determining the size and shape of the nanoparticles. A recent report by Zahoor et al. also demonstrated that amino acids like glutamine, aspartic, and tyrosine are size and shape-controlling agents during silver nanoparticle synthesis [121][42]. Like silver nanoparticles, the copper nanoparticles synthesized using the glucosides isoquercetin and cassinopin reduced the biofilm formation of MRSA with an effective biofilm inhibitory concentration as low as 1 µg/mL. The prepared nanoparticles exhibited antibiofilm activity against MRSA by altering its cell membrane permeability and surface hydrophobicity [122][43].4.1.2. Nanoencapsulation of Natural Compounds with Antibiofilm Activity
Arming natural compounds through the nanoencapsulation procedure has several advantages over surface functionalization techniques. First, they enable sustained release of the bioactive compounds, where the drug release profile is governed by the properties of the polymer being used for encapsulation, such as drug–matrix interactions, solubility, diffusion, and biodegradation [134][44]. Through nanoencapsulation, it is possible to regulate the delivery of natural bioactives to the target site using site-specific stimulants, such as ultrasound, enzymes, pH, and magnetic fields [135,136,137,138][45][46][47][48]. This functional property aids researchers in exploiting nanocarriers as a portable detection system for microbial pathogens to avoid or reduce infectious disease outbreaks [139][49]. Next, nanocarriers enhance the natural compounds’ bioavailability and bioactivity by minimizing the nanoparticles’ size, surface modification, and encapsulating the natural compounds with different polymers [140][50]. Apart from the targeted drug delivery and improving the potential of the encapsulating drug, nanocarriers also protect the encapsulating natural bioactives from degradation, oxidation, and aggregation, which is considered an essential advantage in overseeing the use of nanocarriers over nanoparticles with surface fabrication [141][51].Polymeric Nanoparticles
Polymeric nanoparticles are distinguished by their qualities customized for a specific payload and to the appropriate size, specific cellular trafficking, and easy regulation of drug delivery via improved material engineering. These polymeric materials are either natural (polysaccharides such as chitosan, cellulose, dextran, and others, or polypeptides and proteins, such as albumin, gelatin, legumin, and others) or synthetic (polyglycolide, polycaprolactone, derivatives of polyacrylic acid, poly(ethylene glycol), polylactides (PLA) and copolymers including polylactide co-glycolide (PLGA), and others) [143][52]. There are several reports on the potential of biopolymeric nanosystems against foodborne pathogens’ growth and biofilm formation. These nanoparticles act via electrostatic interactions with the negatively charged EPS in the outer layer of the biofilms. Starch, cellulose, chitosan, cyclodextrin, alginate, and guar gum are a few examples of biopolymers often used to encapsulate natural products. Among these biopolymers, chitosan is a naturally occurring amino polysaccharide polymer with intrinsic antibacterial and antibiofilm properties. It has been a common biopolymer exploited to deliver natural products [144][53]. Earlier reports demonstrated that chitosan is a good choice of biopolymer for encapsulating natural compounds [145,146][54][55] and essential oils [147,148,149][56][57][58] against the biofilm formation of different foodborne pathogens. The use of chitosan nanoparticles is increasing in the active packaging sectors. In this light, a recent work by Khan et al. [150][59] reported that chitosan nanoparticles loaded with usnic acid, which is said to have antibiofilm activity against S. aureus, is efficient in eradicating the biofilm-residing persister cells of foodborne pathogens like S. aureus, E. coli, and L. monocytogenes [151][60]. Next to chitosan, another polysaccharide-based biopolymer that has been much exploited in drug delivery is cyclodextrin. Although three forms of cyclodextrins (α-, β-, and γ-cyclodextrins) are available, β-cyclodextrin is the most frequently used for encapsulation. Cyclodextrins are useful for encapsulating hydrophobic molecules, especially essential oils, as it contains both hydrophilic (outer) and hydrophobic (inner) parts [152][61].Lipid Nanoparticles
Lipid nanoparticles can be categorized as liposomes, nanoemulsions, solid lipid nanoparticles, and nanostructured lipid carriers. Due to their elastic physicochemical properties, well-established safety profiles, and ease of scaling up processes, lipid nanoparticles are some of the most promising tools for the targeted delivery of drugs [165][62]. Most lipids used as nanocarriers are approved by US Food and Drug Administration. Hence, the use of lipid nanoparticles is more prevalent than the previously described metallic and polymeric nanoparticles. Specifically, polyethylene glycol-grafted liposomes were in intelligent mode, which extended their presence for a longer time in the blood circulation without activating phagocytosis-mediated clearance or showing any toxicity [166][63]. Because of the above qualities, lipid-based nanoparticles have been extensively studied for targeting bacterial infections, especially against biofilm formation of bacterial pathogens [167][64]. In this regard, a recent study with nanostructured nano lipid carriers encapsulating olibanum oil was shown to kill sessile C. jejuni cells significantly more than the free oil [168][65]. Because of their industrial scale-up, biocompatibility, low toxicity, and ability to entrap both lipophilic and hydrophilic actives, liposomes are the most studied lipid-based nanoformulations [169][66]. Liposomes are spherical phospholipid bilayer vesicles. Liposomes preferentially adsorb onto biofilm surfaces and penetrate the EPS to prevent bacterial growth [170][67]. In addition to the advantages of using nanocarriers mentioned above, liposomes offer much more peculiar and advanced features, such as protecting volatile and chemically unstable antimicrobial or antibiofilm drugs (e.g., essential oils) against active component loss in air, light, and high temperature conditions during production, storage, and administration [171][68]. Specifically, soy lecithin and cholesterol (5:1)-derived liposomes extended cinnamon oil’s stability to 96 h and increased its antibiofilm activity against MRSA by almost ten times [171][68]. In light of developing drug resistance, liposomes entering the biofilm hide the drug from biofilm inhabitants until they burst. Thereby, the liposome prevents cells from recognizing the drug and developing resistance via horizontal gene transfer [172,173][69][70].Silica Nanoparticles
Similar to other nanoparticles, silica nanoparticles can also encapsulate drugs. Often mesoporous nanoparticles are seen with the encapsulation of active leads after incorporating compounds with stimuli-based release properties [177][71]. Mesoporous silica nanoparticles are adaptable because pore size, particle size, and surface area can be easily modified. It also provides the prolonged release of encapsulated pharmaceuticals over hours or days [178][72]. In a study, mesoporous silica nanoparticles encapsulating different essential oils, such as eucalyptus, orange, and cinnamon, were employed to alter the adhesion, biofilm development, and preformed biofilms of S. aureus and E. coli [179][73]. The activity of the essential oil-encapsulated nanoparticles was found to be dependent on the phytochemicals used for encapsulation.5. Strategies to Enhance Biofilm Clearance
Compared to free compounds, NPs exhibit increased efficacy in preventing biofilm formation and eliminating mature biofilms. In general, antibiofilm activity can be achieved by modifying the surface properties of the nanoparticles, which target bacterial adherence and eliminate the adhering bacterial pathogens. Although the complete eradication of mature biofilms is challenging, it has been accomplished in rare instances. It is believed that at most moments, the cargo was deactivated either by the harbored enzyme or the microenvironment. With antibiotics loaded in lipid liquid crystal nanoparticles, 100 percent biofilm eradication was observed, although a 3-fold greater concentration of free antibiotic failed to eliminate the preformed biofilms [193][74]. In contrast, a few nanoparticles were ineffective. For instance, nanoparticles showed MIC values at the microgram level or failed to eradicate preformed biofilms or kill the biofilm residents [145,191,194][54][75][76]. Zein nanoparticles loaded with anacardic acid could not kill and destroy the residents of preformed biofilms [195][77]. Similarly, liposomal-loaded antilisterial peptide showed a 10-fold increased MIC and antibiofilm inhibition compared to the free peptide [185][78]. This was attributed either to the low drug delivery inside the biofilm or to the inability of the nanoparticles to penetrate the biofilm matrix. Hence, it is crucial to determine appropriate methods to boost NPs’ antibiofilm abilities.5.1. Size and Surface Modifications
The capacity of nanoparticles to penetrate biofilms is directly related to their size. Depending upon the density of the EPS matrix, water channels and meshes in biofilms vary in size from 10 nm to hundreds of nm [196][79]. The diffusion rate of nanoparticles within a biofilm is also directly proportional to its size [197,198][80][81]. It is suggested that nanoparticles with an average size of 130 nm or smaller can penetrate the biofilm matrix and have better biofilm-eradicating potential [199][82]. However, most nanoparticles with this cutoff are attributed to the metallic nanoparticles and necessitate the reassessment of those nanoparticles with small sizes for biofilm penetration and eradication. Notably, the nanoparticles with this cutoff are primarily associated with the metallic ones Similar to size, surface properties of the nanoparticles, like surface charge and hydrophobicity, are also essential factors determining their biofilm penetrating potential. Coating PLA nanoparticles with the positively charged peptide poly-L-lysine enhanced biofilm-eradicating capacity compared to uncharged nanoparticles [164][83]. The cationic polymer chitosan is predicted to prevent bacterial biofilm formation by reacting electrostatically with the negatively charged EPS, proteins, and DNA that make up biofilms [144,200][53][84].5.2. Stimuli-Responsive Release
In the presence of external cues, it is possible to create nanoparticles that trigger the release of encapsulated drugs. An attraction-luring, intelligent, and potential therapeutic technique for optimized drug release is using stimuli-responsive drug delivery systems that are sensitive to a wide range of endogenous stimuli including pH, redox state, and temperature. In this light, the acidic pH of the biofilm can be used as the external stimulus to trigger the release of the encapsulated drug. Cationic farnesol-loaded nanoparticles were synthesized using the co-polymers 2-(dimethylamino) ethyl methacrylate, butyl methacrylate, and 2-propylacrylic acid [205][85]. Similar to pH, the enzymes secreted by bacteria can also be an external cue that can be used to trigger drug release. Specifically, hyaluronidase [207][86], lipase [208][87], gelatinase [209][88], and glutamyl endonuclease [209][88] have also been used as enzyme cues for triggered drug release. Wang and Shukla synthesized an antibiotic nanoparticle loaded on a gelatin core and coated with chitosan and hyaluronic acid [207][86]. Hyaluronidase produced by the bacteria first degraded the hyaluronic acid, exposing the chitosan coating, which in turn aids enhanced biofilm adhesion and penetration. As the chitosan layer expands, more gelatinases could reach the inner core, leading to gelatin breakdown and drug release [206][89]. Compared to the free drug, the drug loaded in the nanoparticle coated with hyaluronic acid enhanced the eradication of the biofilm. As a very affordable and accessible external stimulus, light is widely used for highly precise, controlled drug release from responsive nanocarriers [214][90]. In particular, light in the near-infrared region is appropriate for treating biofilm infections because of its longer wavelength, greater tissue penetration, and low toxicity. In addition, near-infrared region light may operate as a thermal trigger since photothermal agents can convert it to heat. This conversion is often utilized in materials that respond to light since light is considerably simpler to regulate than heat [214][90]. Like near-infrared light, magnetic fields are also widely used in medicine. Magnetic nanoparticles can target particular biofilm infection sites and penetrate protective biofilm matrices with the energy from external magnetic fields. In addition, they have been shown to induce local heat and mechanical stresses, which have the potential to deteriorate polymeric materials and result in drug release.5.3. Combined Strategies
The combination of two different antibiofilm strategies will have synergistic activity by enhancing biofilm eradication. The innovative nanoencapsulation approach of liposome co-encapsulating antibiotics with metals improves their antibacterial and antibiofilm properties. For instance, gallium targets iron metabolism and iron-dependent cellular processes to impede bacterial growth and biofilm formation [217,218][91][92]. Following this approach, the growth and biofilm formation of P. aeruginosa was targeted by DPPC/dipalmitoyl phosphatidylglycerol (DPPG) liposomes containing gallium and gentamicin. This formulation outperformed liposome-loaded gentamicin and the free antibiotic against planktonic and biofilm P. aeruginosa cells. This metal–antibiotic-loaded liposome also eliminated bacterial biofilms and QS signaling and lowered gallium toxicity [219][93]. Bismuth and bismuth-thiol, like gallium, alter iron uptake, alginate expression, lipopolysaccharides, virulence factors, bacterial adhesion, and biofilm formation to inhibit a broad spectrum of microorganisms [220,221,222,223][94][95][96][97]. Antimicrobial photodynamic therapy (aPDT) eliminates specific cells by inducing reactive oxygen species. This approach utilizes three different factors such as light (at an appropriate wavelength), a photosensitizer (a compound activated by light), and molecular oxygen (for the generation of ROS) [228,229][98][99]. Singlet oxygen, produced explicitly by this approach, triggers cell death by interacting with practically all cellular components and biomolecules. Among several synthetic photosensitizers, natural compounds such as curcumin, hypericin, and flavin derivatives, are used [229][99].References
- Kostakioti, M.; Hadjifrangiskou, M.; Hultgren, S.J. Bacterial biofilms: Development, dispersal, and therapeutic strategies in the dawn of the postantibiotic era. Cold Spring Harb. Perspect. Med. 2013, 3, a010306.
- Vestby, L.K.; Grønseth, T.; Simm, R.; Nesse, L.L. Bacterial biofilm and its role in the pathogenesis of disease. Antibiotics 2020, 9, 59.
- Arunachalam, K.; Davoodbasha, M. Imaging Bacteria and Biofilm by Field Emission Scanning Electron Microscopy. In Analytical Methodologies for Biofilm Research; Springer Protocols Handbooks; Springer: New York, NY, USA, 2021; pp. 205–222.
- van Wolferen, M.; Orell, A.; Albers, S.-V. Archaeal biofilm formation. Nat. Rev. Microbiol. 2018, 16, 699–713.
- Cámara, M.; Green, W.; MacPhee, C.E.; Rakowska, P.D.; Raval, R.; Richardson, M.C.; Slater-Jefferies, J.; Steventon, K.; Webb, J.S. Microbiomes. Economic significance of biofilms: A multidisciplinary and cross-sectoral challenge. Npj Biofilms Microbiomes 2022, 8, 42.
- Alvarez-Ordóñez, A.; Coughlan, L.M.; Briandet, R.; Cotter, P.D. Biofilms in food processing environments: Challenges and opportunities. Annu. Rev. Food Sci. Technol. 2019, 10, 173–195.
- Galie, S.; García-Gutiérrez, C.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Biofilms in the food industry: Health aspects and control methods. Front. Microbiol. 2018, 9, 898.
- Rodríguez-Campos, D.; Rodríguez-Melcón, C.; Alonso-Calleja, C.; Capita, R. Persistent Listeria monocytogenes isolates from a poultry-processing facility form more biofilm but do not have a greater resistance to disinfectants than sporadic strains. Pathogens 2019, 8, 250.
- Cadena, M.; Kelman, T.; Marco, M.L.; Pitesky, M.J.F. Understanding antimicrobial resistance (AMR) profiles of Salmonella biofilm and planktonic bacteria challenged with disinfectants commonly used during poultry processing. Foods 2019, 8, 275.
- Pang, X.; Song, X.; Chen, M.; Tian, S.; Lu, Z.; Sun, J.; Li, X.; Lu, Y.; Yuk, H.-G. Combating biofilms of foodborne pathogens with bacteriocins by lactic acid bacteria in the food industry. Compr. Rev. Food Sci. Food Saf. 2022, 21, 1657–1676.
- Uddin Mahamud, A.G.M.S.; Nahar, S.; Ashrafudoulla, M.; Park, S.H.; Ha, S.-D. Insights into antibiofilm mechanisms of phytochemicals: Prospects in the food industry. Crit. Rev. Food Sci. Nutr. 2022.
- Deng, Y.; Liu, Y.; Li, J.; Wang, X.; He, S.; Yan, X.; Shi, Y.; Zhang, W.; Ding, L. Marine natural products and their synthetic analogs as promising antibiofilm agents for antibiotics discovery and development. Eur. J. Med. Chem. 2022, 239, 114513.
- Martinengo, P.; Arunachalam, K.; Shi, C.J.F. Polyphenolic Antibacterials for Food Preservation: Review, Challenges, and Current Applications. Foods 2021, 10, 2469.
- Chitlapilly Dass, S.; Wang, R. Biofilm through the Looking Glass: A Microbial Food Safety Perspective. Pathogens 2022, 11, 346.
- Srey, S.; Jahid, I.K.; Ha, S.-D. Biofilm formation in food industries: A food safety concern. Food Control 2013, 31, 572–585.
- Lin, Y.; Briandet, R.; Kovács, Á.T. Bacillus cereus sensu lato biofilm formation and its ecological importance. Biofilm 2022, 4, 100070.
- Liu, X.-Y.; Hu, Q.; Xu, F.; Ding, S.-Y.; Zhu, K. Characterization of Bacillus cereus in dairy products in China. Toxins 2020, 12, 454.
- Carter, M.Q.; Feng, D.; Li, H.H. Curli fimbriae confer shiga toxin-producing Escherichia coli a competitive trait in mixed biofilms. Food Microbiol. 2019, 82, 482–488.
- Yang, S.-C.; Lin, C.-H.; Aljuffali, I.A.; Fang, J.-Y. Current pathogenic Escherichia coli foodborne outbreak cases and therapy development. Arch. Microbiol. 2017, 199, 811–825.
- Olaimat, A.N.; Al-Holy, M.A.; Shahbaz, H.M.; Al-Nabulsi, A.A.; Abu Ghoush, M.H.; Osaili, T.M.; Ayyash, M.M.; Holley, R.A. Emergence of antibiotic resistance in Listeria monocytogenes isolated from food products: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1277–1292.
- Arteaga, V.; Lamas, A.; Regal, P.; Vázquez, B.; Miranda, J.M.; Cepeda, A.; Franco, C.M. Antimicrobial activity of apitoxin from Apis mellifera in Salmonella enterica strains isolated from poultry and its effects on motility, biofilm formation and gene expression. Microb. Pathog. 2019, 137, 103771.
- Srinivasan, R.; Kannappan, A.; Shi, C.; Lin, X. Marine bacterial secondary metabolites: A treasure house for structurally unique and effective antimicrobial compounds. Mar. Drugs 2021, 19, 530.
- Camargo, A.C.; de Paula, O.A.; Todorov, S.D.; Nero, L.A. In vitro evaluation of bacteriocins activity against Listeria monocytogenes biofilm formation. Appl. Biochem. Biotechnol. 2016, 178, 1239–1251.
- Abdelhadi, I.M.A.; Sofy, A.R.; Hmed, A.A.; Refaey, E.E.; Soweha, H.E.; Abbas, M.A. Discovery of Polyvalent Myovirus (vB_STM-2) Phage as a natural antimicrobial system to lysis and biofilm removal of Salmonella Typhimurium Isolates from various food sources. Sustainability 2021, 13, 11602.
- Martínez-Graciá, C.; González-Bermúdez, C.A.; Cabellero-Valcárcel, A.M.; Santaella-Pascual, M.; Frontela-Saseta, C. Use of herbs and spices for food preservation: Advantages and limitations. Curr. Opin. Food Sci. 2015, 6, 38–43.
- Nazzaro, F.; Fratianni, F.; Coppola, R. Quorum sensing and phytochemicals. Int. J. Mol. Sci. 2013, 14, 12607–12619.
- Chen, Y.; Liu, T.; Wang, K.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein inhibits Staphylococcus aureus biofilm formation and the quorum sensing system in vitro. PLoS ONE 2016, 11, e0153468.
- Shu, Q.; Lou, H.; Wei, T.; Liu, X.; Chen, Q. Contributions of Glycolipid Biosurfactants and Glycolipid-Modified Materials to Antimicrobial Strategy: A Review. Pharmaceutics 2021, 13, 227.
- Silveira, V.A.I.; Nishio, E.K.; Freitas, C.A.U.Q.; Amador, I.R.; Kobayashi, R.K.T.; Caretta, T.; Macedo, F.; Celligoi, M.A.P. Production and antimicrobial activity of sophorolipid against Clostridium perfringens and Campylobacter jejuni and their additive interaction with lactic acid. Biocatal. Agric. Biotechnol. 2019, 21, 101287.
- Zhang, X.; Ashby, R.; Solaiman, D.K.Y.; Uknalis, J.; Fan, X. Inactivation of Salmonella spp. and Listeria spp. by palmitic, stearic, and oleic acid sophorolipids and thiamine dilauryl sulfate. Front. Microbiol. 2016, 7, 2076.
- Elgamoudi, B.A.; Korolik, V. Campylobacter Biofilms: Potential of Natural Compounds to Disrupt Campylobacter jejuni Transmission. Int. J. Mol. Sci. 2021, 22, 12159.
- Hintz, T.; Matthews, K.K.; Di, R. The Use of Plant Antimicrobial Compounds for Food Preservation. BioMed Res. Int. 2015, 2015, 246264.
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta (BBA) Biomembr. 1993, 1147, 132–136.
- Driessen, A.J.M.; van den Hooven, H.W.; Kuiper, W.; Van de Camp, M.; Sahl, H.-G.; Konings, R.N.H.; Konings, W.N. Mechanistic studies of lantibiotic-induced permeabilization of phospholipid vesicles. Biochemistry 1995, 34, 1606–1614.
- Brewer, M.S. Natural Antioxidants: Sources, Compounds, Mechanisms of Action, and Potential Applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247.
- Alvarez, M.; Moreira, M.d.R.; Roura, S.; Ayala-Zavala, J.; González-Aguilar, G.A. Using Natural Antimicrobials to Enhance the Safety and Quality of Fresh and Processed Fruits and Vegetables: TYPES of Antimicrobials. In Handbook of Natural Antimicrobials for Food Safety and Quality; Woodhead Publishing: Cambridge, UK, 2015; pp. 287–313.
- Zhang, L.; Gu, F.X.; Chan, J.M.; Wang, A.Z.; Langer, R.S.; Farokhzad, O.C. Nanoparticles in medicine: Therapeutic applications and developments. Clin. Pharmacol. Ther. 2008, 83, 761–769.
- Slavin, Y.N.; Asnis, J.; Häfeli, U.O.; Bach, H. Metal nanoparticles: Understanding the mechanisms behind antibacterial activity. J. Nanobiotechnol. 2017, 15, 65.
- Barros, C.H.N.; Devlin, H.; Hiebner, D.W.; Vitale, S.; Quinn, L.; Casey, E. Enhancing curcumin’s solubility and antibiofilm activity via silica surface modification. Nanoscale Adv. 2020, 2, 1694–1708.
- Menon, S.; Rajeshkumar, S.; Kumar, V. A review on biogenic synthesis of gold nanoparticles, characterization, and its applications. Resour. Effic. Technol. 2017, 3, 516–527.
- Sharma, D.; Kanchi, S.; Bisetty, K. Biogenic synthesis of nanoparticles: A review. Arab. J. Chem. 2019, 12, 3576–3600.
- Zahoor, M.; Nazir, N.; Iftikhar, M.; Naz, S.; Zekker, I.; Burlakovs, J.; Uddin, F.; Kamran, A.W.; Kallistova, A.; Pimenov, N.; et al. A review on silver nanoparticles: Classification, various methods of synthesis, and their potential roles in biomedical applications and water treatment. Water 2021, 13, 2216.
- Lotha, R.; Shamprasad, B.R.; Sundaramoorthy, N.S.; Nagarajan, S.; Sivasubramanian, A. Biogenic phytochemicals (cassinopin and isoquercetin) capped copper nanoparticles (ISQ/) inhibits MRSA biofilms. Microb. Pathog. 2019, 132, 178–187.
- Begines, B.; Ortiz, T.; Pérez-Aranda, M.; Martínez, G.; Merinero, M.; Argüelles-Arias, F.; Alcudia, A. Polymeric nanoparticles for drug delivery: Recent developments and future prospects. Nanomaterials 2020, 10, 1403.
- Avramović, N.; Mandić, B.; Savić-Radojević, A.; Simić, T. Polymeric nanocarriers of drug delivery systems in cancer therapy. Pharmaceutics 2020, 12, 298.
- Elbialy, N.S.; Aboushoushah, S.F.; Sofi, B.F.; Noorwali, A. Multifunctional curcumin-loaded mesoporous silica nanoparticles for cancer chemoprevention and therapy. Microporous Mesoporous Mater. 2020, 291, 109540.
- Di, J.; Kim, J.; Hu, Q.; Jiang, X.; Gu, Z. Spatiotemporal drug delivery using laser-generated-focused ultrasound system. J. Control. Release 2015, 220, 592–599.
- Sirivisoot, S.; Harrison, B.S. Magnetically stimulated ciprofloxacin release from polymeric microspheres entrapping iron oxide nanoparticles. Int. J. Nanomed. 2015, 10, 4447–4458.
- Wang, Z.; Chen, Z.; Gao, N.; Ren, J.; Qu, X. Transmutation of personal glucose meters into portable and highly sensitive microbial pathogen detection platform. Small 2015, 11, 4970–4975.
- Gunasekaran, T.; Haile, T.; Nigusse, T.; Dhanaraju, M.D. Nanotechnology: An effective tool for enhancing bioavailability and bioactivity of phytomedicine. Asian Pac. J. Trop. Biomed. 2014, 4, S1–S7.
- Pircalabioru, G.G.; Chifiriuc, M.-C. Nanoparticulate drug-delivery systems for fighting microbial biofilms: From bench to bedside. Future Microbiol. 2020, 15, 679–698.
- Michalak, G.; Głuszek, K.; Piktel, E.; Deptuła, P.; Puszkarz, I.; Niemirowicz, K.; Bucki, R. Polymeric nanoparticles—A novel solution for delivery of antimicrobial agents. Med. Stud. 2016, 32, 56–62.
- Perinelli, D.R.; Fagioli, L.; Campana, R.; Lam, J.K.W.; Baffone, W.; Palmieri, G.F.; Casettari, L.; Bonacucina, G. Chitosan-based nanosystems and their exploited antimicrobial activity. Eur. J. Pharm. Sci. 2018, 117, 8–20.
- Siddhardha, B.; Pandey, U.; Kaviyarasu, K.; Pala, R.; Syed, A.; Bahkali, A.H.; Elgorban, A.M. Chrysin-loaded chitosan nanoparticles potentiates antibiofilm activity against Staphylococcus aureus. Pathogens 2020, 9, 115.
- Xu, J.; Lin, Q.; Sheng, M.; Ding, T.; Li, B.; Gao, Y.; Tan, Y. Antibiofilm effect of cinnamaldehyde-chitosan nanoparticles against the biofilm of Staphylococcus aureus. Antibiotics 2022, 11, 1403.
- Liu, T.; Liu, L. Fabrication and characterization of chitosan nanoemulsions loading thymol or thyme essential oil for the preservation of refrigerated pork. Int. J. Biol. Macromol. 2020, 162, 1509–1515.
- Granata, G.; Stracquadanio, S.; Leonardi, M.; Napoli, E.; Malandrino, G.; Cafiso, V.; Stefani, S.; Geraci, C. Oregano and thyme essential oils encapsulated in chitosan nanoparticles as effective antimicrobial agents against foodborne pathogens. Molecules 2021, 26, 4055.
- Cai, M.; Wang, Y.; Wang, R.; Li, M.; Zhang, W.; Yu, J.; Hua, R. Antibacterial and antibiofilm activities of chitosan nanoparticles loaded with Ocimum basilicum L. essential oil. Int. J. Biol. Macromol. 2022, 202, 122–129.
- Khan, F.; Yu, H.; Kim, Y.-M. Bactericidal activity of usnic acid-chitosan nanoparticles against persister cells of biofilm-forming pathogenic bacteria. Mar. Drugs 2020, 18, 270.
- Pompilio, A.; Riviello, A.; Crocetta, V.; Giuseppe, F.D.; Pomponio, S.; Sulpizio, M.; Ilio, C.D.; Angelucci, S.; Barone, L.; Giulio, A.D.; et al. Evaluation of antibacterial and antibiofilm mechanisms by usnic acid against methicillin-resistant Staphylococcus aureus. Future Microbiol. 2016, 11, 1315–1338.
- Folch-Cano, C.; Yazdani-Pedram, M.; Olea-Azar, C. Inclusion and functionalization of polymers with cyclodextrins: Current applications and future prospects. Molecules 2014, 19, 14066–14079.
- del Pozo-Rodríguez, A.; Solinís, M.Á.; Rodríguez-Gascón, A. Applications of lipid nanoparticles in gene therapy. Eur. J. Pharm. Biopharm. 2016, 109, 184–193.
- Ahmed, K.; Muiruri, P.W.; Jones, G.H.; Scott, M.J.; Jones, M.N. The effect of grafted poly(ethylene glycol) on the electrophoretic properties of phospholipid liposomes and their adsorption to bacterial biofilms. Colloids Surf. A Physicochem. Eng. Asp. 2001, 194, 287–296.
- Wang, D.-Y.; van der Mei, H.C.; Ren, Y.; Busscher, H.J.; Shi, L. Lipid-based antimicrobial delivery-systems for the treatment of bacterial infections. Front. Chem. 2020, 7, 872.
- Ribeiro, L.N.d.M.; de Paula, E.; Rossi, D.A.; Martins, F.A.; de Melo, R.T.; Monteiro, G.P.; Breitkreitz, M.C.; Goulart, L.R.; Fonseca, B.B. Nanocarriers from natural lipids with in vitro activity against Campylobacter jejuni. Front. Cell. Infect. Microbiol. 2021, 10, 571040.
- Filipczak, N.; Pan, J.; Yalamarty, S.S.K.; Torchilin, V.P. Recent advancements in liposome technology. Adv. Drug Deliv. Rev. 2020, 156, 4–22.
- Wang, Y. Liposome as a delivery system for the treatment of biofilm-mediated infections. J. Appl. Microbiol. 2021, 131, 2626–2639.
- Cui, H.; Li, W.; Li, C.; Vittayapadung, S.; Lin, L. Liposome containing cinnamon oil with antibacterial activity against methicillin-resistant Staphylococcus aureus biofilm. Biofouling 2016, 32, 215–225.
- Dos Santos Ramos, M.A.; Da Silva, P.B.; Spósito, L.; De Toledo, L.G.; Bonifácio, B.V.; Rodero, C.F.; Dos Santos, K.C.; Chorilli, M.; Bauab, T.M. Nanotechnology-based drug delivery systems for control of microbial biofilms: A review. Int. J. Nanomed. 2018, 13, 1179–1213.
- Rukavina, Z.; Vanić, Ž. Current Trends in Development of Liposomes for Targeting Bacterial Biofilms. Pharmaceutics 2016, 8, 18.
- Wu, Y.; Long, Y.; Li, Q.-L.; Han, S.; Ma, J.; Yang, Y.-W.; Gao, H. Layer-by-layer (LBL) self-assembled biohybrid nanomaterials for efficient antibacterial applications. ACS Appl. Mater. Interfaces 2015, 7, 17255–17263.
- Tang, F.; Li, L.; Chen, D. Mesoporous silica nanoparticles: Synthesis, biocompatibility and drug delivery. Adv. Mater. 2012, 24, 1504–1534.
- Balaure, P.C.; Boarca, B.; Popescu, R.C.; Savu, D.; Trusca, R.; Vasile, B.Ș.; Grumezescu, A.M.; Holban, A.M.; Bolocan, A.; Andronescu, E. Bioactive mesoporous silica nanostructures with anti-microbial and anti-biofilm properties. Int. J. Pharm. 2017, 531, 35–46.
- Thorn, C.R.; Carvalho-Wodarz, C.d.S.; Horstmann, J.C.; Lehr, C.-M.; Prestidge, C.A.; Thomas, N. Tobramycin liquid crystal nanoparticles eradicate cystic fibrosis-related Pseudomonas aeruginosa biofilms. Small 2021, 17, 2100531.
- Nosrati, M.; Ranjbar, R. Investigation of the antibacterial and biofilm inhibitory activities of Prangos acaulis (DC.) Bornm in nanoparticulated formulation. Nanotechnology 2022, 33, 385103.
- Dalcin, A.J.F.; Santos, C.G.; Gündel, S.S.; Roggia, I.; Raffin, R.P.; Ourique, A.F.; Santos, R.C.V.; Gomes, P. Anti biofilm effect of dihydromyricetin-loaded nanocapsules on urinary catheter infected by Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2017, 156, 282–291.
- Lima, R.A.; de Souza, S.L.X.; Lima, L.A.; Batista, A.L.X.; de Araújo, J.T.C.; Sousa, F.F.O.; Rolim, J.; Bandeira, T. Antimicrobial effect of anacardic acid-loaded zein nanoparticles loaded on Streptococcus mutans biofilms. Braz. J. Microbiol. 2020, 51, 1623–1630.
- Pu, C.; Tang, W. The antibacterial and antibiofilm efficacies of a liposomal peptide originating from rice bran protein against Listeria monocytogenes. Food Funct. 2017, 8, 4159–4169.
- Liu, Y.; Shi, L.; Su, L.; van der Mei, H.C.; Jutte, P.C.; Ren, Y.; Busscher, H.J. Nanotechnology-based antimicrobials and delivery systems for biofilm-infection control. Chem. Soc. Rev. 2019, 48, 428–446.
- Peulen, T.O.; Wilkinson, K.J. Diffusion of nanoparticles in a biofilm. Environ. Sci. Technol. 2011, 45, 3367–3373.
- MubarakAli, D.; Arunkumar, J.; Pooja, P.; Subramanian, G.; Thajuddin, N.; Alharbi, N.S. Synthesis and characterization of biocompatibility of tenorite nanoparticles and potential property against biofilm formation. Saudi Pharm. J. 2015, 23, 421–428.
- Forier, K.; Messiaen, A.-S.; Raemdonck, K.; Nelis, H.; De Smedt, S.; Demeester, J.; Coenye, T.; Braeckmans, K. Probing the size limit for nanomedicine penetration into Burkholderia multivorans and Pseudomonas aeruginosa biofilms. J. Control. Release 2014, 195, 21–28.
- Da Costa, D.; Exbrayat-Héritier, C.; Rambaud, B.; Megy, S.; Terreux, R.; Verrier, B.; Primard, C. Surface charge modulation of rifampicin-loaded PLA nanoparticles to improve antibiotic delivery in Staphylococcus aureus biofilms. J. Nanobiotechnol. 2021, 19, 12.
- Jiang, F.; Deng, Y.; Yeh, C.-K.; Sun, Y. Quaternized chitosans bind onto preexisting biofilms and eradicate pre-attached microorganisms. J. Mater. Chem. B 2014, 2, 8518–8527.
- Horev, B.; Klein, M.I.; Hwang, G.; Li, Y.; Kim, D.; Koo, H.; Benoit, D.S. pH-activated nanoparticles for controlled topical delivery of farnesol to disrupt oral biofilm virulence. ACS Nano 2015, 9, 2390–2404.
- Wang, Y.; Shukla, A. Bacteria-responsive biopolymer-coated nanoparticles for biofilm penetration and eradication. Biomater. Sci. 2022, 10, 2831–2843.
- Liu, Y.; Ren, Y.; Li, Y.; Su, L.; Zhang, Y.; Huang, F.; Liu, J.; Liu, J.; van Kooten, T.G.; An, Y.; et al. Nanocarriers with conjugated antimicrobials to eradicate pathogenic biofilms evaluated in murine in vivo and human ex vivo infection models. Acta Biomater. 2018, 79, 331–343.
- Ding, Y.; Hao, Y.; Yuan, Z.; Tao, B.; Chen, M.; Lin, C.; Liu, P.; Cai, K. A dual-functional implant with an enzyme-responsive effect for bacterial infection therapy and tissue regeneration. Biomater. Sci. 2020, 8, 1840–1854.
- Sims, K.R.; Liu, Y.; Hwang, G.; Jung, H.I.; Koo, H.; Benoit, D.S.W. Enhanced design and formulation of nanoparticles for anti-biofilm drug delivery. Nanoscale 2018, 11, 219–236.
- Ren, Y.; Liu, H.; Liu, X.; Zheng, Y.; Li, Z.; Li, C.; Yeung, K.W.K.; Zhu, S.; Liang, Y.; Cui, Z.; et al. Photoresponsive Materials for Antibacterial Applications. Cell Rep. Phys. Sci. 2020, 1, 100245.
- Kaneko, Y.; Thoendel, M.; Olakanmi, O.; Britigan, B.E.; Singh, P.K. The transition metal gallium disrupts Pseudomonas aeruginosa iron metabolism and has antimicrobial and antibiofilm activity. J. Clin. Investig. 2007, 117, 877–888.
- Olakanmi, O.; Britigan, B.E.; Schlesinger, L.S. Gallium disrupts iron metabolism of mycobacteria residing within human macrophages. Infect. Immun. 2000, 68, 5619–5627.
- Halwani, M.; Yebio, B.; Suntres, Z.E.; Alipour, M.; Azghani, A.O.; Omri, A. Co-encapsulation of gallium with gentamicin in liposomes enhances antimicrobial activity of gentamicin against Pseudomonas aeruginosa. J. Antimicrob. Chemother. 2008, 62, 1291–1297.
- Domenico, P.; Baldassarri, L.; Schoch, P.E.; Kaehler, K.; Sasatsu, M.; Cunha, B.A. Activities of bismuth thiols against staphylococci and staphylococcal biofilms. Antimicrob. Agents Chemother. 2001, 45, 1417–1421.
- Domenico, P.; Reich, J.; Madonia, W.; Cunha, B.A. Resistance to bismuth among Gram-negative bacteria is dependent upon iron and its uptake. J. Antimicrob. Chemother. 1996, 38, 1031–1040.
- Huang, C.T.; Stewart, P.S. Reduction of polysaccharide production in Pseudomonas aeruginosa biofilms by bismuth dimercaprol (BisBAL) treatment. J. Antimicrob. Chemother. 1999, 44, 601–605.
- Folsom, J.P.; Baker, B.; Stewart, P.S. In vitro efficacy of bismuth thiols against biofilms formed by bacteria isolated from human chronic wounds. J. Appl. Microbiol. 2011, 111, 989–996.
- de Annunzio, S.R.; de Freitas, L.M.; Blanco, A.L.; da Costa, M.M.; Carmona-Vargas, C.C.; de Oliveira, K.T.; Fontana, C.R. Susceptibility of Enterococcus faecalis and Propionibacterium acnes to antimicrobial photodynamic therapy. J. Photochem. Photobiol. B Biol. 2018, 178, 545–550.
- Silva, A.F.; de Oliveira Silva, J.V.; Campanerut-Sá, P.A.Z.; Mikcha, J.M.G. Chapter 13—Photoinactivation of biofilms. In Recent Trends in Biofilm Science and Technology; Simoes, M., Borges, A., Chaves Simoes, L., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 295–306.