Tauopathies are a class of neurodegenerative diseases, including Alzheimer’s disease, and are characterized by intraneuronal tau inclusion in the brain and the patient’s cognitive decline with obscure pathogenesis. Heparan sulfate proteoglycans, a major type of extracellular matrix, have been believed to involve in tauopathies. The heparan sulfate proteoglycans co-deposit with tau in Alzheimer’s patient brain, directly bind to tau and modulate tau secretion, internalization, and aggregation.
- tauopathy
- heparan sulfate proteoglycan
- tau
- Alzheimer’s disease
1. Introduction
2. The Tau Protein
Tau protein belongs to the microtubule-associated proteins family [4]. Tau is found predominantly in the axon of the adult neurons and at low levels in the cell body, dendritic spines, and axonal presynaptic terminal [4]. The subcellular distribution of the tau is developmentally and environmentally regulated and isoform-dependent [5,6,7,8][5][6][7][8]. In addition, tau is detected at low levels in glial cells and outside cells [2,7,9][2][7][9]. The various subcellular, extracellular, and cell-type expressions indicate that tau may play various functions under different circumstances. Human tau is encoded by a single gene, microtubule-associated protein tau (Mapt), located on chromosome 17q21. Mapt gene comprises 16 exons (Figure 1. By alternative splicing of exon 2 (E2), exon 3 (E3), and exon 10 (E10), six isoforms of tau are expressed in the normal adult human brain [2]. The largest isoform contains 441 amino acid residues, including an N-terminus projection domain with two inserts (N1 and N2), a proline-rich domain, and a microtubule-binding repeat domain composed of four repeat motifs (R1–R4) that mediate microtubule-binding and tau aggregation [2]. The isoforms differ in the absence or presence of N1, N2, and R2 domains leading to the following 6 isoforms: 2N4R, 1N4R, 0N4R, 2N3R, 1N3R, and 0N3R (Figure 1) [1,3,10][1][3][10]. The N1 and N2 are encoded by E2 and E3, respectively, and R2 is encoded by E10. Besides the six tau isoforms, researchers also widely use another two truncated tau proteins, the K18 and K19, which contain only the four and three microtubule-binding domains, respectively (Figure 1). Given the differential distribution of tau in different cell compartments and presence of various tau isoforms, tau likely plays different functions in different environments.
3. Tau in Physiological States
As a microtubule-associated protein, Tau promotes axonal outgrowth through stabilizing neuronal microtubules [2]. Early studies indeed showed that tau stabilizes the axonal microtubules, promotes the assembly of microtubules, and regulates the dynamic instability of the microtubules (Figure 2A) [11[11][12][13],12,13], suggesting that tau is critical for developing a healthy neuron. However, knockout of mouse Mapt, the gene encoding tau protein, does not lead to a severe developmental defect or overt abnormalities at young ages [14,15,16][14][15][16]. Additionally, in neuronal cell line studies, tau was shown to inhibit axonal transport through multiple mechanisms, including competing with kinesin or dynein for binding to microtubules [17], competing with other cargos for binding to kinesin [18[18][19],19], reducing the number of cargo-associated kinesin motors [20], and releasing cargos from the kinesin chains [21] (Figure 2A). However, knockout or overexpression of tau does not alter axonal transport in cultured primary neurons [22,23,24][22][23][24]. These observations suggest that tau might be dispensable for microtubule assembly, stability, and axonal transport. The lack of the expected microtubule and axonal transport defects in the Mapt−/− mice is likely due to redundancy in function among tau and compensatory effect from other microtubule-associated proteins for the loss of tau [15,25][15][25]. Tau has been suggested to regulate synaptic physiology and plasticity, although it is expressed at a low level in the dendrites (Figure 2A) [26,27,28][26][27][28]. Tetsuya et al. observed a selective deficit in long-term depression (LTD) but not long-term potential (LTP) in Mapt−/− mice [27]. A study reported by Ahmed et al. observed a severe deficit in LTP, but no change in LTD, in another Mapt−/− mouse line [28]. Tau is also expressed in the nuclei and is thought to maintain the integrity of the genomic DNA (Figure 2A) [29]. Recent studies have emerged that tau may act generally as a scaffolding protein interacting with various kinases and phosphatases to regulate phosphorylation-based signaling pathways, functioning as a signaling hub protein within cells [30].
4. Tau in Pathological States
Physiological tau is a natively unfolded and highly soluble protein with little tendency for aggregation [9]. Under pathogenic conditions, the dynamics and equilibrium of tau-microtubule binding are disrupted, leading to tau aggregation and oligomerization into paired helical filaments (PHFs) and, further, the neurofibrillary tangles (NFTs) which accumulate in neurons, glia, and extracellular space (Figure 2B) [2]. The formation of tau aggregates is more strongly correlated with cognitive decline than the distribution of senile plaques formed by amyloid beta (Aβ) protein deposits, another pathological hallmark of AD [41][31].5. Heparan Sulfate Proteoglycans
Heparan sulfate proteoglycans (HSPGs) are macromolecules ubiquitously expressed in mammalian tissues. They are comprised of a core protein to which one or more HS glycosaminoglycan (GAG) chains are covalently attached [87,88][32][33]. The HSPGs are classified based on the location of their core proteins (Figure 3A). Syndecans (SDCs) and glypicans are the two major membrane-bound PGs that are linked to the plasma membrane by a transmembrane domain or a glycosylphosphatidylinositol (GPI) linker, respectively. In addition, there are three “part-time” cell-surface PGs, betaglycans, neuropilins, and CD44v3, which do not always have an HS chain moiety and are located on the cell surface through their transmembrane domains. Other HSPGs include agrin, perlecan, type XVIII collagen in the extracellular matrix (ECM), and serglycin in the intracellular secretory vesicles (Figure 3A) [87,88][32][33].
6. Heparan Sulfate-Tau Interaction: The Related Structures
HS and heparin, a highly sulfated form of HS, directly bind to tau protein [97,98][39][40]. Snow et al. and Su et al. observed by ultrastructural immunolocalization that HS co-localizes with NFTs in neurons in the brains of AD patients [99[41][42],100], suggesting that HS interacts with tau in the AD brain. Like other heparin-protein interactions [101[43][44],102], the tau binds HS and the binding is mediated by electrostatic interaction through the highly positively charged residues/domain within the tau protein and the highly negatively charged sulfate residues within HS. Further studies determined that the hexapeptides 275VQIINK280 in R2 and 306VQIVYK311 in R3 of tau bind to HS [103,104,105,106,107][45][46][47][48][49]. In NMR titration of heparin to K18, L282 displayed the most chemical shift perturbation, suggesting that the two lysines near 275VQIINK280 in R2, K280 and K281, are likely crucial to heparin and HS binding. A recent study further showed that Arixtra, a sulfated pentamer analog of heparin, bound proline-rich region II (PRR2) with 4 times stronger binding affinity than R2 [108][50]. These studies indicate that the major HS binding sites within tau protein are in the R2 and PRR2 domains (Figure 4A).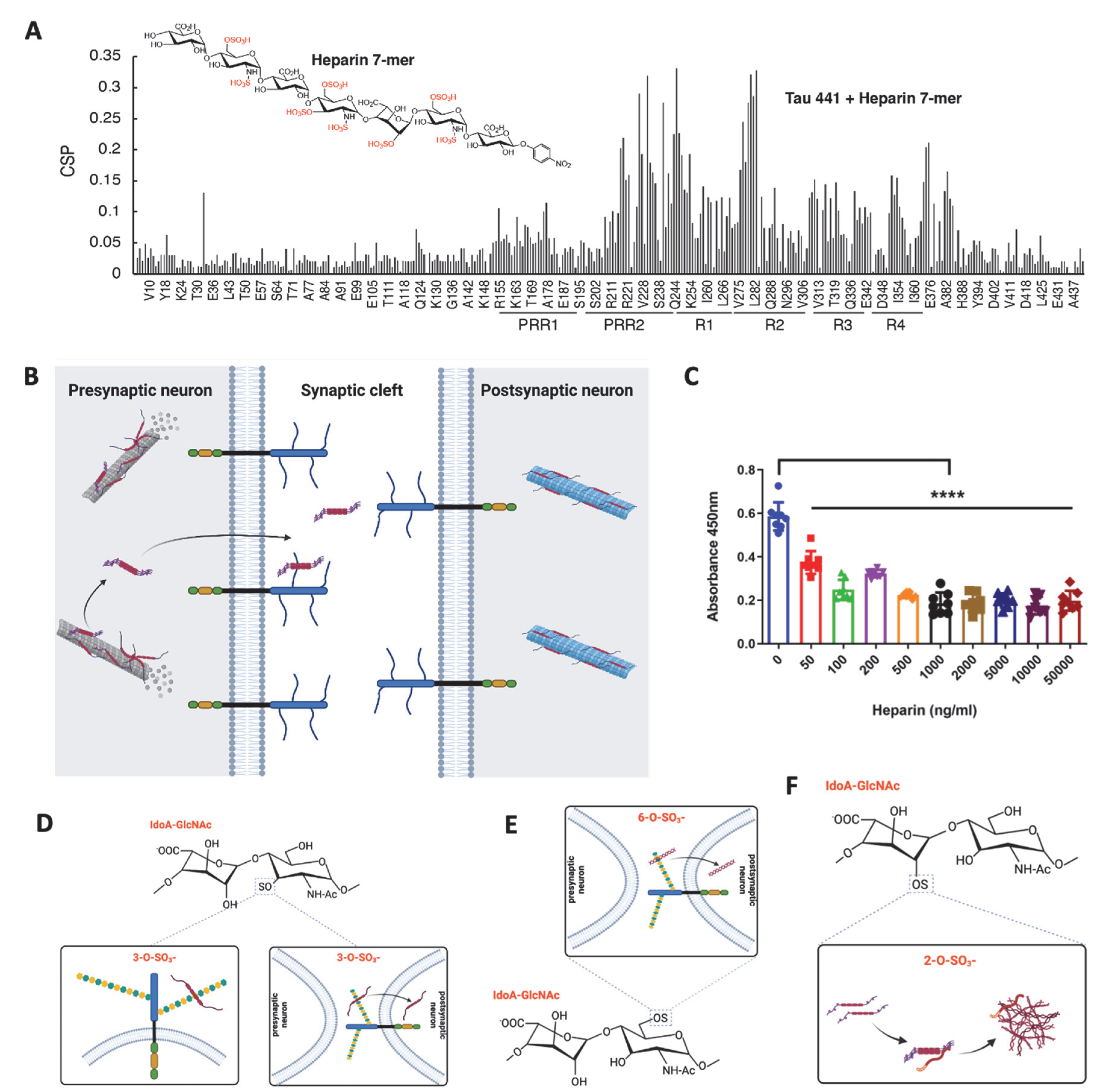
7. The Role of Heparan Sulfate in the Tau-Mediated Pathological Process
HS has been suggested to play an essential role in each stage of the prion-like propagation of tau pathology, including Tau secretion, cell surface binding, internalization, and aggregation [111][51].7.1. HS in Tau Secretion
Tau is predominately an intracellular protein and has also been found in the extracellular space under both physiological and pathological conditions [112][52]. Tau is continuously secreted under physiological conditions without cell death, indicating some functional roles of the extracellular tau [113,114,115][53][54][55]. Several studies have demonstrated that tau does not follow the conventional secretory pathway but uses multiple unconventional secretory pathways [116,117][56][57]. Merezhko et al. showed that phosphorylated, oligomeric tau clusters on the plasma membrane in neuronal cells and is secreted in the vesicle-free form in an unconventional process, and the secretion was supported by cell surface HSPGs, possibly by facilitating its release after membrane penetration [118][58]. Katsinelos et al. further delineated that in the cytosol, free tau interacts with phosphatidylinositol 4,5-bisphosphate enriched at the inner leaflet of the plasma membrane, leading to its translocation across the plasma membrane via HSPGs (Figure 4B) [119][59].7.2. HS in Tau Cell Surface Binding
The association between tau and the cell surface implicates tau protein uptake and related intracellular signaling. Tau was observed to bind to endothelial cell surface and the cell surface binding was inhibited by externally added heparin (Figure 4C), suggesting that HS mediates the cell surface binding of tau, which is consistent with other groups’ findings in which HSPGs mediated the binding of tau to C17.2 cells [111,120][51][60]. In addition, knockout of HS 3-O-sulfotransferase-1 (Hs3st1) reduces 3-O-sulfation of HS and attenuated endothelial cell surface HS-mediated tau protein binding, supporting the high-affinity tau binding site within HS containing 3-O-sulfate (Figure 4D) [93,110][35][61].7.3. HS in Tau Internalization
In 2013, Holmes et al. first demonstrated that HSPGs is a critical mediator for tau uptake in mouse neural progenitor cell line [111][51]. Using differently sized and chemically modified heparins, the same group further determined that tau aggregates required a somewhat specific HS architecture with defined sulfate moieties containing N- and 6-O-sulfations, confirmed by reduced tau cellular uptake upon knockout of Ndst1 or Hs6st2, the genes responsible for N-sulfation and 6-O-sulfation modification in HS biosynthesis, respectively [95][37]. Rauch et al. also reported that tau protein internalization depends on 6-O-sulfation of HS (Figure 4E) [121][62]. The 3-O-sulfation has been proven to enhance HS-mediated tau internalization. Sepulveda-Diaz et al. reported that 3-O-sulfated HS could be internalized into cells where HS interacts with tau and promotes tau phosphorylation [110][61]. In the recent study, the researchers showed that introducing a 3-O-sulfate significantly increased the binding of an HS dodecasaccharide to tau and knockout of Hs3st1, which reduces about 50% 3-O-sulfation of HS in mouse lung endothelial cells [122][63], significantly decreased tau uptake by the cells (Figure 4D) [93][35]. Knockout of Hs3st1 in HEK293T cells appeared not to affect tau uptake [95][37], which might be due to the low expression of Hs3st1 in this cell line (https://www.proteinatlas.org/ENSG00000002587-HS3ST1/cell+line (accessed on 20 November 2022)). These studies demonstrated that HS facilitates tau protein internalization and further support that the tau-binding HS structure contains N-, 6-O-, and 3-O-sulfations, in agreement with previous biochemical binding studies [95][37]. In contrast, HSPGs were dispensable for tau protein uptake by primary astrocytes, revealing that HS’s function in facilitating tau internalization is cell-type dependent [95,123][37][64].7.4. HS in Tau Aggregation
HS was found to accumulate with NFTs in the AD brain, suggesting that HS may promote tau aggregation in the brain [100][42]. Arrasate et al. incubated the isolated PHFs from AD patients with heparinase and found that the PHF morphology was changed after digestion [124][65]. These observations suggested that HS may facilitate tau protein aggregation to exacerbate tauopathy. This hypothesis has been supported by the regular in vitro tau aggregation experimental setting, which requires the addition of polyanionic cofactors such as RNA or heparin to initiate the aggregation [125,126,127,128,129,130][66][67][68][69][70][71]. This facilitation depends on the direct binding of heparin to soluble tau monomers, which is thought to neutralize the positive, repulsive charges on tau and allow more contact between monomers [125,126,130][66][67][71]. Townsend et al. examined truncated tau (Δtau187, residue 255–441) aggregation induced with chemically modified heparins [127][68]. Removal of 6-O-sulfation, not 2-O-sulfation, reduces heparin’s binding affinity for Δtau187, which in agreement with other study findings, showing 6-O-sulfation is required for HS to bind tau protein [95,106][37][48]. However, tau aggregation is considerably slower in the presence of 2-O-desulfated heparin than with N- or 6-O-desulfated heparin, indicating that 2-O-sulfation contributes more than 6-O and N-sulfation in facilitating tau aggregation (Figure 4F), apparently due to 2-O-sulfation promoting tau primary and secondary nucleation and filament elongation [127][68]. However, the sulfation pattern dependence has not been determined in the aggregation of full-length tau and phosphorylated tau. It may be different, especially considering the importance of PRR2 in tau binding. In addition, Sepulveda-Diaz, J.E. et al. reported that Hs3st2 increases tau phosphorylation in a cell model and a zebrafish model of tauopathy, showing that HS promotes tau phosphorylation to facilitate tau aggregation indirectly. This is supported by Huynh et al. reporting that Hs3st2 expression induces the cell-autonomous oligomerization of tau and the phosphorylation of tau in an in vitro cell model [131][72].8. Aberrant HSPG Expression in AD and Other Tauopathies
The proteoglycan cores of HSPGs are differentially expressed in AD compared with normal human brains, which has been shown to impact tau internalization [132][73]. The impact of Alzheimer’s disease on membrane-bound HSPG agrin distribution has been documented since 1999; in normal brains, agrin is in the soluble fraction of detergent extracted samples, while in AD brains, agrin shifts to the insoluble fraction [133][74]. One study found that genes associated with syndecan-4 (SDC4), followed by SDC3, SDC2, agrin, and serglycin (the dominant intracellular PG in immune cells) [134][75] were consistently overexpressed in AD brains compared with controls [135][76]. SDC3 has the highest affinity for tau monomer, but interestingly, overexpression of SDC3 and -4 in K562 cells (lymphoblast cells with low endogenous HS) did not lead to a greater internalization of tau, but increased fibrillation and accumulation of fibrils on the cell surface [132][73]. These studies emphasize the impact of tauopathies on the specific core proteins of HSPGs. Several studies have documented altered HS expression in AD patients (Table 1). Su et al. examined 7 AD patients and 4 age-matched controls and observed that the number and intensity of the HS co-staining with PHFs were denser in AD than in control brains [99][41]. Shimizu et al. examined 25 AD patients with 10 non-demented elderly patient controls. They detected a 9.3-fold HS increase in the hippocampus and a 6.6-fold HS increase in gyrus frontalis superior in the AD patients. They also observed that HS is most abundantly expressed in the basement membrane of capillary endothelial cells [136][77]. Other groups’ studies confirmed the abnormal HS expression in the AD brain [92,137][34][78].Clinical Diagnosis | Predominant Tau Isoforms | Human Brain Samples | GAGs/Gene Expression in Disease | GAGs Function in Disease | Reference | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
AD | 3R + 4R Tau | 7 AD vs. 4 control | HS ↑ | N/A |
[99] |
[41] |
|||||||||||
AD | 3R + 4R Tau | N/A | N/A | Helicity of PHFs changed (potential) |
[124] |
[65] |
|||||||||||
AD | 3R + 4R Tau | 25 AD vs. 10 control | HS ↑ | N/A |
[136] |
[77] |
|||||||||||
AD | 3R + 4R Tau | 20 AD vs. 20 control | Sulf1 -; Sulf2 ↓ | N/A |
[138] |
[79] |
|||||||||||
AD | 3R + 4R Tau | 5 AD vs. 5 control | HS ↑; Ndst2 ↑; Hs3st2 ↑; Hs3st4 ↑; Glce ↑; HPSE ↑ | HS-tau binding capacity ↑ |
[92] |
[34] |
|||||||||||
AD | 3R + 4R Tau | 18 AD vs. 6 control | Altered expression of multiple HS biosynthesis/remodeling genes | N/A |
[139] |
[80] |
|||||||||||
AD | 3R + 4R Tau | 5 AD vs. 5 control | HS ↑; 3-o-sulfation ↑ | N/A |
[137] |
[78] |
References
- Zhang, Y.; Wu, K.M.; Yang, L.; Dong, Q.; Yu, J.T. Tauopathies: New perspectives and challenges. Mol. Neurodegener. 2022, 17, 28.
- Wang, Y.; Mandelkow, E. Tau in physiology and pathology. Nat. Rev. Neurosci. 2016, 17, 5–21.
- Gotz, J.; Halliday, G.; Nisbet, R.M. Molecular Pathogenesis of the Tauopathies. Annu. Rev. Pathol. 2019, 14, 239–261.
- Ittner, A.; Ittner, L.M. Dendritic Tau in Alzheimer’s Disease. Neuron. 2018, 99, 13–27.
- Drubin, D.G.; Caput, D.; Kirschner, M.W. Studies on the expression of the microtubule-associated protein, tau, during mouse brain development, with newly isolated complementary DNA probes. J. Cell Biol. 1984, 98, 1090–1097.
- Sultan, A.; Nesslany, F.; Violet, M.; Bégard, S.; Loyens, A.; Talahari, S.; Mansuroglu, Z.; Marzin, D.; Sergeant, N.; Humez, S.; et al. Nuclear tau, a key player in neuronal DNA protection. J. Biol. Chem. 2011, 286, 4566–4575.
- Papasozomenos, S.C.; Binder, L.I. Phosphorylation determines two distinct species of Tau in the central nervous system. Cell Motil. Cytoskeleton 1987, 8, 210–226.
- Liu, C.; Götz, J. Profiling murine tau with 0N, 1N and 2N isoform-specific antibodies in brain and peripheral organs reveals distinct subcellular localization, with the 1N isoform being enriched in the nucleus. PLoS ONE 2013, 8, e84849.
- Chang, C.W.; Shao, E.; Mucke, L. Tau: Enabler of diverse brain disorders and target of rapidly evolving therapeutic strategies. Science 2021, 371, eabb8255.
- Andreadis, A. Tau gene alternative splicing: Expression patterns, regulation and modulation of function in normal brain and neurodegenerative diseases. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2005, 1739, 91–103.
- Drubin, D.G.; Kirschner, M.W. Tau protein function in living cells. J. Cell Biol. 1986, 103, 2739–2746.
- Panda, D.; Goode, B.L.; Feinstein, S.C.; Wilson, L. Kinetic stabilization of microtubule dynamics at steady state by tau and microtubule-binding domains of tau. Biochemistry 1995, 34, 11117–11127.
- Trinczek, B.; Biernat, J.; Baumann, K.; Mandelkow, E.M.; Mandelkow, E. Domains of tau protein, differential phosphorylation, and dynamic instability of microtubules. Mol. Biol. Cell 1995, 6, 1887–1902.
- Dawson, H.N.; Ferreira, A.; Eyster, M.V.; Ghoshal, N.; Binder, L.I.; Vitek, M.P. Inhibition of neuronal maturation in primary hippocampal neurons from tau deficient mice. J. Cell Sci. 2001, 114, 1179–1187.
- Harada, A.; Oguchi, K.; Okabe, S.; Kuno, J.; Terada, S.; Ohshima, T.; Sato-Yoshitake, R.; Takei, Y.; Noda, T.; Hirokawa, N. Altered microtubule organization in small-calibre axons of mice lacking tau protein. Nature 1994, 369, 488–491.
- Tucker, K.L.; Meyer, M.; Barde, Y.A. Neurotrophins are required for nerve growth during development. Nat. Neurosci. 2001, 4, 29–37.
- Dixit, R.; Ross, J.L.; Goldman, Y.E.; Holzbaur, E.L. Differential regulation of dynein and kinesin motor proteins by tau. Science 2008, 319, 1086–1089.
- Konzack, S.; Thies, E.; Marx, A.; Mandelkow, E.M.; Mandelkow, E. Swimming against the tide: Mobility of the microtubule-associated protein tau in neurons. J. Neurosci. 2007, 27, 9916–9927.
- Utton, M.A.; Noble, W.J.; Hill, J.E.; Anderton, B.H.; Hanger, D.P. Molecular motors implicated in the axonal transport of tau and alpha-synuclein. J. Cell Sci. 2005, 118, 4645–4654.
- Vershinin, M.; Carter, B.C.; Razafsky, D.S.; King, S.J.; Gross, S.P. Multiple-motor based transport and its regulation by Tau. Proc. Natl. Acad. Sci. USA 2007, 104, 87–92.
- Kanaan, N.M.; Morfini, G.A.; LaPointe, N.E.; Pigino, G.F.; Patterson, K.R.; Song, Y.; Andreadis, A.; Fu, Y.; Brady, S.T.; Binder, L.I. Pathogenic forms of tau inhibit kinesin-dependent axonal transport through a mechanism involving activation of axonal phosphotransferases. J. Neurosci. 2011, 31, 9858–9868.
- Vossel, K.A.; Xu, J.C.; Fomenko, V.; Miyamoto, T.; Suberbielle, E.; Knox, J.A.; Ho, K.; Kim, D.H.; Yu, G.Q.; Mucke, L. Tau reduction prevents Aβ-induced axonal transport deficits by blocking activation of GSK3β. J. Cell Biol. 2015, 209, 419–433.
- Vossel, K.A.; Zhang, K.; Brodbeck, J.; Daub, A.C.; Sharma, P.; Finkbeiner, S.; Cui, B.; Mucke, L. Tau reduction prevents Abeta-induced defects in axonal transport. Science 2010, 330, 198.
- Yuan, A.; Kumar, A.; Peterhoff, C.; Duff, K.; Nixon, R.A. Axonal transport rates in vivo are unaffected by tau deletion or overexpression in mice. J. Neurosci. 2008, 28, 1682–1687.
- Takei, Y.; Teng, J.; Harada, A.; Hirokawa, N. Defects in axonal elongation and neuronal migration in mice with disrupted tau and map1b genes. J. Cell Biol. 2000, 150, 989–1000.
- Frandemiche, M.L.; De Seranno, S.; Rush, T.; Borel, E.; Elie, A.; Arnal, I.; Lanté, F.; Buisson, A. Activity-dependent tau protein translocation to excitatory synapse is disrupted by exposure to amyloid-beta oligomers. J. Neurosci. 2014, 34, 6084–6097.
- Kimura, T.; Whitcomb, D.J.; Jo, J.; Regan, P.; Piers, T.; Heo, S.; Brown, C.; Hashikawa, T.; Murayama, M.; Seok, H.; et al. Microtubule-associated protein tau is essential for long-term depression in the hippocampus. Philos. Trans. R Soc. Lond B. Biol. Sci. 2014, 369, 20130144.
- Ahmed, T.; Van der Jeugd, A.; Blum, D.; Galas, M.C.; D’Hooge, R.; Buee, L.; Balschun, D. Cognition and hippocampal synaptic plasticity in mice with a homozygous tau deletion. Neurobiol. Aging 2014, 35, 2474–2478.
- Violet, M.; Delattre, L.; Tardivel, M.; Sultan, A.; Chauderlier, A.; Caillierez, R.; Talahari, S.; Nesslany, F.; Lefebvre, B.; Bonnefoy, E.; et al. A major role for Tau in neuronal DNA and RNA protection in vivo under physiological and hyperthermic conditions. Front. Cell Neurosci. 2014, 8, 84.
- Mueller, R.L.; Combs, B.; Alhadidy, M.M.; Brady, S.T.; Morfini, G.A.; Kanaan, N.M. Tau: A Signaling Hub Protein. Front. Mol. Neurosci. 2021, 14, 647054.
- Bierer, L.M.; Hof, P.R.; Purohit, D.P.; Carlin, L.; Schmeidler, J.; Davis, K.L.; Perl, D.P. Neocortical neurofibrillary tangles correlate with dementia severity in Alzheimer’s disease. Arch. Neurol. 1995, 52, 81–88.
- Sarrazin, S.; Lamanna, W.C.; Esko, J.D. Heparan sulfate proteoglycans. Cold Spring Harb. Perspect. Biol. 2011, 3.
- Bernfield, M.; Gotte, M.; Park, P.W.; Reizes, O.; Fitzgerald, M.L.; Lincecum, J.; Zako, M. Functions of cell surface heparan sulfate proteoglycans. Annu. Rev. Biochem. 1999, 68, 729–777.
- Huynh, M.B.; Ouidja, M.O.; Chantepie, S.; Carpentier, G.; Maiza, A.; Zhang, G.; Vilares, J.; Raisman-Vozari, R.; Papy-Garcia, D. Glycosaminoglycans from Alzheimer’s disease hippocampus have altered capacities to bind and regulate growth factors activities and to bind tau. PLoS ONE 2019, 14, e0209573.
- Zhao, J.; Zhu, Y.; Song, X.; Xiao, Y.; Su, G.; Liu, X.; Wang, Z.; Xu, Y.; Liu, J.; Eliezer, D.; et al. 3-O-Sulfation of Heparan Sulfate Enhances Tau Interaction and Cellular Uptake. Angew. Chem. Int. Ed. Engl. 2020, 59, 1818–1827.
- Snow, A.D.; Cummings, J.A.; Lake, T. The Unifying Hypothesis of Alzheimer’s Disease: Heparan Sulfate Proteoglycans/Glycosaminoglycans Are Key as First Hypothesized Over 30 Years Ago. Front. Aging Neurosci. 2021, 13, 710683.
- Stopschinski, B.E.; Holmes, B.B.; Miller, G.M.; Manon, V.A.; Vaquer-Alicea, J.; Prueitt, W.L.; Hsieh-Wilson, L.C.; Diamond, M.I. Specific glycosaminoglycan chain length and sulfation patterns are required for cell uptake of tau versus alpha-synuclein and beta-amyloid aggregates. J. Biol. Chem. 2018, 293, 10826–10840.
- Ledin, J.; Staatz, W.; Li, J.P.; Gotte, M.; Selleck, S.; Kjellen, L.; Spillmann, D. Heparan sulfate structure in mice with genetically modified heparan sulfate production. J. Biol. Chem. 2004, 279, 42732–42741.
- Mah, D.; Zhao, J.; Liu, X.; Zhang, F.; Liu, J.; Wang, L.; Linhardt, R.; Wang, C. The Sulfation Code of Tauopathies: Heparan Sulfate Proteoglycans in the Prion Like Spread of Tau Pathology. Front. Mol. Biosci. 2021, 8, 671458.
- Alavi Naini, S.M.; Soussi-Yanicostas, N. Heparan Sulfate as a Therapeutic Target in Tauopathies: Insights From Zebrafish. Front. Cell Dev. Biol. 2018, 6, 163.
- Su, J.H.; Cummings, B.J.; Cotman, C.W. Localization of heparan sulfate glycosaminoglycan and proteoglycan core protein in aged brain and Alzheimer’s disease. Neuroscience 1992, 51, 801–813.
- Snow, A.D.; Mar, H.; Nochlin, D.; Sekiguchi, R.T.; Kimata, K.; Koike, Y.; Wight, T.N. Early accumulation of heparan sulfate in neurons and in the beta-amyloid protein-containing lesions of Alzheimer’s disease and Down’s syndrome. Am. J. Pathol. 1990, 137, 1253–1270.
- Farshi, P.; Ohlig, S.; Pickhinke, U.; Hoing, S.; Jochmann, K.; Lawrence, R.; Dreier, R.; Dierker, T.; Grobe, K. Dual roles of the Cardin-Weintraub motif in multimeric Sonic hedgehog. J. Biol. Chem. 2011, 286, 23608–23619.
- Torrent, M.; Nogues, M.V.; Andreu, D.; Boix, E. The “CPC clip motif”: A conserved structural signature for heparin-binding proteins. PLoS ONE 2012, 7, e42692.
- Kato, T.; Sasaki, H.; Katagiri, T.; Sasaki, H.; Koiwai, K.; Youki, H.; Totsuka, S.; Ishii, T. The binding of basic fibroblast growth factor to Alzheimer’s neurofibrillary tangles and senile plaques. Neurosci. Lett. 1991, 122, 33–36.
- Perry, G.; Siedlak, S.L.; Richey, P.; Kawai, M.; Cras, P.; Kalaria, R.N.; Galloway, P.G.; Scardina, J.M.; Cordell, B.; Greenberg, B.D.; et al. Association of heparan sulfate proteoglycan with the neurofibrillary tangles of Alzheimer’s disease. J. Neurosci. 1991, 11, 3679–3683.
- Spillantini, M.G.; Tolnay, M.; Love, S.; Goedert, M. Microtubule-associated protein tau, heparan sulphate and alpha-synuclein in several neurodegenerative diseases with dementia. Acta Neuropathol. 1999, 97, 585–594.
- Zhao, J.; Huvent, I.; Lippens, G.; Eliezer, D.; Zhang, A.; Li, Q.; Tessier, P.; Linhardt, R.J.; Zhang, F.; Wang, C. Glycan Determinants of Heparin-Tau Interaction. Biophys. J. 2017, 112, 921–932.
- Mukrasch, M.D.; Biernat, J.; von Bergen, M.; Griesinger, C.; Mandelkow, E.; Zweckstetter, M. Sites of tau important for aggregation populate -structure and bind to microtubules and polyanions. J. Biol. Chem. 2005, 280, 24978–24986.
- Murray, A.Y.L.; Gibson, J.M.; Liu, J.; Eliezer, D.; Lippens, G.; Zhang, F.; Linhardt, R.J.; Zhao, J.; Wang, C. Proline-Rich Region II (PRR2) Plays an Important Role in Tau-Glycan Interaction: An NMR Study. Biomolecules 2022, 12, 1573.
- Holmes, B.B.; DeVos, S.L.; Kfoury, N.; Li, M.; Jacks, R.; Yanamandra, K.; Ouidja, M.O.; Brodsky, F.M.; Marasa, J.; Bagchi, D.P.; et al. Heparan sulfate proteoglycans mediate internalization and propagation of specific proteopathic seeds. Proc. Natl. Acad. Sci. USA 2013, 110, E3138–E3147.
- Pérez, M.; Avila, J.; Hernández, F. Propagation of Tau via Extracellular Vesicles. Front. Neurosci. 2019, 13, 698.
- Toledo, J.B.; Zetterberg, H.; van Harten, A.C.; Glodzik, L.; Martinez-Lage, P.; Bocchio-Chiavetto, L.; Rami, L.; Hansson, O.; Sperling, R.; Engelborghs, S.; et al. Alzheimer’s disease cerebrospinal fluid biomarker in cognitively normal subjects. Brain 2015, 138, 2701–2715.
- Sutphen, C.L.; Jasielec, M.S.; Shah, A.R.; Macy, E.M.; Xiong, C.; Vlassenko, A.G.; Benzinger, T.L.; Stoops, E.E.; Vanderstichele, H.M.; Brix, B.; et al. Longitudinal Cerebrospinal Fluid Biomarker Changes in Preclinical Alzheimer Disease During Middle Age. JAMA Neurol. 2015, 72, 1029–1042.
- Pilliod, J.; Desjardins, A.; Pernegre, C.; Jamann, H.; Larochelle, C.; Fon, E.A.; Leclerc, N. Clearance of intracellular tau protein from neuronal cells via VAMP8-induced secretion. J. Biol. Chem. 2020, 295, 17827–17841.
- Merezhko, M.; Uronen, R.L.; Huttunen, H.J. The Cell Biology of Tau Secretion. Front. Mol. Neurosci. 2020, 13, 569818.
- Xu, Y.; Cui, L.; Dibello, A.; Wang, L.; Lee, J.; Saidi, L.; Lee, J.G.; Ye, Y. DNAJC5 facilitates USP19-dependent unconventional secretion of misfolded cytosolic proteins. Cell Discov. 2018, 4, 11.
- Merezhko, M.; Brunello, C.A.; Yan, X.; Vihinen, H.; Jokitalo, E.; Uronen, R.L.; Huttunen, H.J. Secretion of Tau via an Unconventional Non-vesicular Mechanism. Cell Rep. 2018, 25, 2027–2035.e2024.
- Katsinelos, T.; Zeitler, M.; Dimou, E.; Karakatsani, A.; Muller, H.M.; Nachman, E.; Steringer, J.P.; Ruiz de Almodovar, C.; Nickel, W.; Jahn, T.R. Unconventional Secretion Mediates the Trans-cellular Spreading of Tau. Cell Rep. 2018, 23, 2039–2055.
- Mirbaha, H.; Holmes, B.B.; Sanders, D.W.; Bieschke, J.; Diamond, M.I. Tau Trimers Are the Minimal Propagation Unit Spontaneously Internalized to Seed Intracellular Aggregation. J. Biol. Chem. 2015, 290, 14893–14903.
- Sepulveda-Diaz, J.E.; Alavi Naini, S.M.; Huynh, M.B.; Ouidja, M.O.; Yanicostas, C.; Chantepie, S.; Villares, J.; Lamari, F.; Jospin, E.; van Kuppevelt, T.H.; et al. HS3ST2 expression is critical for the abnormal phosphorylation of tau in Alzheimer’s disease-related tau pathology. Brain 2015, 138, 1339–1354.
- Rauch, J.N.; Chen, J.J.; Sorum, A.W.; Miller, G.M.; Sharf, T.; See, S.K.; Hsieh-Wilson, L.C.; Kampmann, M.; Kosik, K.S. Tau Internalization is Regulated by 6-O Sulfation on Heparan Sulfate Proteoglycans (HSPGs). Sci. Rep. 2018, 8, 6382.
- Qiu, H.; Shi, S.; Yue, J.; Xin, M.; Nairn, A.V.; Lin, L.; Liu, X.; Li, G.; Archer-Hartmann, S.A.; Dela Rosa, M.; et al. A mutant-cell library for systematic analysis of heparan sulfate structure-function relationships. Nat. Methods 2018, 15, 889–899.
- Perea, J.R.; Lopez, E.; Diez-Ballesteros, J.C.; Avila, J.; Hernandez, F.; Bolos, M. Extracellular Monomeric Tau Is Internalized by Astrocytes. Front. Neurosci. 2019, 13, 442.
- Arrasate, M.; Pérez, M.; Valpuesta, J.M.; Avila, J. Role of glycosaminoglycans in determining the helicity of paired helical filaments. Am. J. Pathol. 1997, 151, 1115–1122.
- Perez, M.; Valpuesta, J.M.; Medina, M.; Montejo de Garcini, E.; Avila, J. Polymerization of tau into filaments in the presence of heparin: The minimal sequence required for tau-tau interaction. J. Neurochem. 1996, 67, 1183–1190.
- Fichou, Y.; Lin, Y.; Rauch, J.N.; Vigers, M.; Zeng, Z.; Srivastava, M.; Keller, T.J.; Freed, J.H.; Kosik, K.S.; Han, S. Cofactors are essential constituents of stable and seeding-active tau fibrils. Proc. Natl. Acad. Sci. USA 2018, 115, 13234–13239.
- Townsend, D.; Fullwood, N.J.; Yates, E.A.; Middleton, D.A. Aggregation Kinetics and Filament Structure of a Tau Fragment Are Influenced by the Sulfation Pattern of the Cofactor Heparin. Biochemistry 2020, 59, 4003–4014.
- Paudel, H.K.; Li, W. Heparin-induced conformational change in microtubule-associated protein Tau as detected by chemical cross-linking and phosphopeptide mapping. J. Biol. Chem. 1999, 274, 8029–8038.
- Martin, L.; Latypova, X.; Wilson, C.M.; Magnaudeix, A.; Perrin, M.-L.; Yardin, C.; Terro, F. Tau protein kinases: Involvement in Alzheimer’s disease. Ageing Res. Rev. 2013, 12, 289–309.
- Maïza, A.; Chantepie, S.; Vera, C.; Fifre, A.; Huynh, M.B.; Stettler, O.; Ouidja, M.O.; Papy-Garcia, D. The role of heparan sulfates in protein aggregation and their potential impact on neurodegeneration. FEBS Lett. 2018, 592, 3806–3818.
- Huynh, M.B.; Rebergue, N.; Merrick, H.; Gomez-Henao, W.; Jospin, E.; Biard, D.S.F.; Papy-Garcia, D. HS3ST2 expression induces the cell autonomous aggregation of tau. Sci. Rep. 2022, 12, 10850.
- Hudak, A.; Kusz, E.; Domonkos, I.; Josvay, K.; Kodamullil, A.T.; Szilak, L.; Hofmann-Apitius, M.; Letoha, T. Contribution of syndecans to cellular uptake and fibrillation of alpha-synuclein and tau. Sci. Rep. 2019, 9, 16543.
- Donahue, J.E.; Berzin, T.M.; Rafii, M.S.; Glass, D.J.; Yancopoulos, G.D.; Fallon, J.R.; Stopa, E.G. Agrin in Alzheimer’s disease: Altered solubility and abnormal distribution within microvasculature and brain parenchyma. Proc. Natl. Acad. Sci. USA 1999, 96, 6468–6472.
- Kolset, S.O.; Pejler, G. Serglycin: A structural and functional chameleon with wide impact on immune cells. J. Immunol. 2011, 187, 4927–4933.
- Lorente-Gea, L.; Garcia, B.; Martin, C.; Ordiales, H.; Garcia-Suarez, O.; Pina-Batista, K.M.; Merayo-Lloves, J.; Quiros, L.M.; Fernandez-Vega, I. Heparan Sulfate Proteoglycans Undergo Differential Expression Alterations in Alzheimer Disease Brains. J. Neuropathol. Exp. Neurol. 2020, 79, 474–483.
- Shimizu, H.; Ghazizadeh, M.; Sato, S.; Oguro, T.; Kawanami, O. Interaction between β-amyloid protein and heparan sulfate proteoglycans from the cerebral capillary basement membrane in Alzheimer’s disease. J. Clin. Neurosci. 2009, 16, 277–282.
- Wang, Z.; Arnold, K.; Dhurandahare, V.M.; Xu, Y.; Pagadala, V.; Labra, E.; Jeske, W.; Fareed, J.; Gearing, M.; Liu, J. Analysis of 3-O-Sulfated Heparan Sulfate Using Isotopically L.Labeled Oligosaccharide Calibrants. Anal. Chem. 2022, 94, 2950–2957.
- Roberts, R.O.; Kang, Y.N.; Hu, C.; Moser, C.D.; Wang, S.; Moore, M.J.; Graham, R.P.; Lai, J.-P.; Petersen, R.C.; Roberts, L.R. Decreased Expression of Sulfatase 2 in the Brains of Alzheimer’s Disease Patients: Implications for Regulation of Neuronal Cell Signaling. J. Alzheimer’s Dis. Rep. 2017, 1, 115–124.
- Pérez-López, N.; Martín, C.; García, B.; Solís-Hernández, M.P.; Rodríguez, D.; Alcalde, I.; Merayo, J.; Fernández-Vega, I.; Quirós, L.M. Alterations in the Expression of the Genes Responsible for the Synthesis of Heparan Sulfate in Brains With Alzheimer Disease. J. Neuropathol. Exp. Neurol. 2021, 80, 446–456.