Neutrophils are surveillance cells, and the first to react and migrate to sites of inflammation and infection following a chemotactic gradient. Neutrophils play a key role in both sterile inflammation and infection, performing a wide variety of effector functions such as degranulation, phagocytosis, ROS production and release of neutrophil extracellular traps (NETs). Healthy term labour requires a sterile pro-inflammatory process, whereas one of the most common causes of spontaneous preterm birth is microbial driven. Peripheral neutrophilia has long been described during pregnancy, and evidence exists demonstrating neutrophils infiltrating the cervix, uterus and foetal membranes during both term and preterm deliveries. Their presence supports a role in tissue remodelling via their effector functions.
- neutrophils
- pregnancy
- term labour
- premature labour
- inflammation
1. Neutrophils and Their Effector Functions
2. Neutrophils during Healthy Pregnancy
Pregnancy involves a complex immunological state that requires a balance between tolerance and immunosuppression to allow growth of the semi-allogeneic foetus, whilst maintaining an effective defence mechanism that can protect against infection [30][13]. There is acknowledgement of a highly active and regulated immune response with shifts between predominance of an anti-inflammatory and pro-inflammatory immune milieu, depending on gestational timepoint and anatomical location [31,32][14][15]. In the most simplistic description, the first trimester is described as a pro-inflammatory phase, to enable implantation. The second phase resembles an anti-inflammatory phase, where a shift in cytokine bias towards T helper 2 phenotype is thought to contribute to foetal growth. Thirdly, a pro-inflammatory switch is seen in preparation for labour, with influx of immune cells into gestational tissue. Recent signatures from peripheral blood immune cells also support a systemic immune clock for human pregnancy [33][16].2.1. Peripheral Blood Neutrophils in Healthy Pregnancy
There is an increase in white blood cell counts in pregnancy, especially towards the end; and a further increase at the time of labour [34][17]. This increase in leukocyte counts is mainly driven by an increase in peripheral granulocyte numbers [35][18], with several studies confirming that pregnancy is associated with a mild neutrophilia [36][19]. The immunophenotype of maternal peripheral neutrophils has been shown to reflect increased activation during pregnancy compared to non-pregnant women. Increased capacity for phagocytosis [37[20][21],38], increased ROS production [4[4][22][23],39,40], and arginase metabolism [41][24] have all been demonstrated in peripheral blood neutrophils of pregnant women compared to non-pregnant controls. Furthermore, response to common pathogens associated with an infectious aetiology of preterm birth is heightened in neutrophils taken from pregnant women compared to non-pregnant controls [42][25].2.2. Neutrophils at the Maternal–Foetal Interface in Healthy Pregnancy
Neutrophils are detected in the decidua of healthy human pregnancy from as early as the first trimester. It is thought that decidual neutrophils play a key role in tissue remodelling and placental vascularization-like spiral artery remodelling [49][26]. Detection of neutrophils in amniotic fluid is also seen and increases with advancing gestational age [50][27]. Their presence is likely to ensure protection against any invading pathogens in a highly sterile environment, to protect the growing foetus. In contrast, there is sparse distribution of neutrophils in myometrium and the cervix prior to the onset of labour [51,52][28][29].3. The Role of Neutrophils in Human Term Labour
Successful pregnancy requires uterine quiescence, the foetus to be contained in the amniotic sac, and the cervix to remain long and closed [53,54][30][31]. The process of parturition requires the onset of uterine contractions, rupture of the foetal membranes, and dilatation of the cervix. This ultimately leads to delivery of the foetus and the placenta, and subsequent involution of the uterus and tissue repair of the cervix. These events are driven by mechanical, inflammatory, and endocrine processes, whose regulation are complex and interlinked [55,56][32][33]. These key processes depend on pro-labour mediators such as COX-2, prostaglandins, matrix metalloproteinases and pro-inflammatory cytokines such as IL-8 [57[34][35][36],58,59], all of which can be produced by neutrophils [60,61,62,63][37][38][39][40]. Despite the triggers of term labour being poorly understood, it is associated with both systemic and local sterile inflammation, with much evidence to support a functional role for neutrophils. Figure 31 illustrates the physiological processes and their anatomical locations where neutrophils are considered to play a key role in term birth.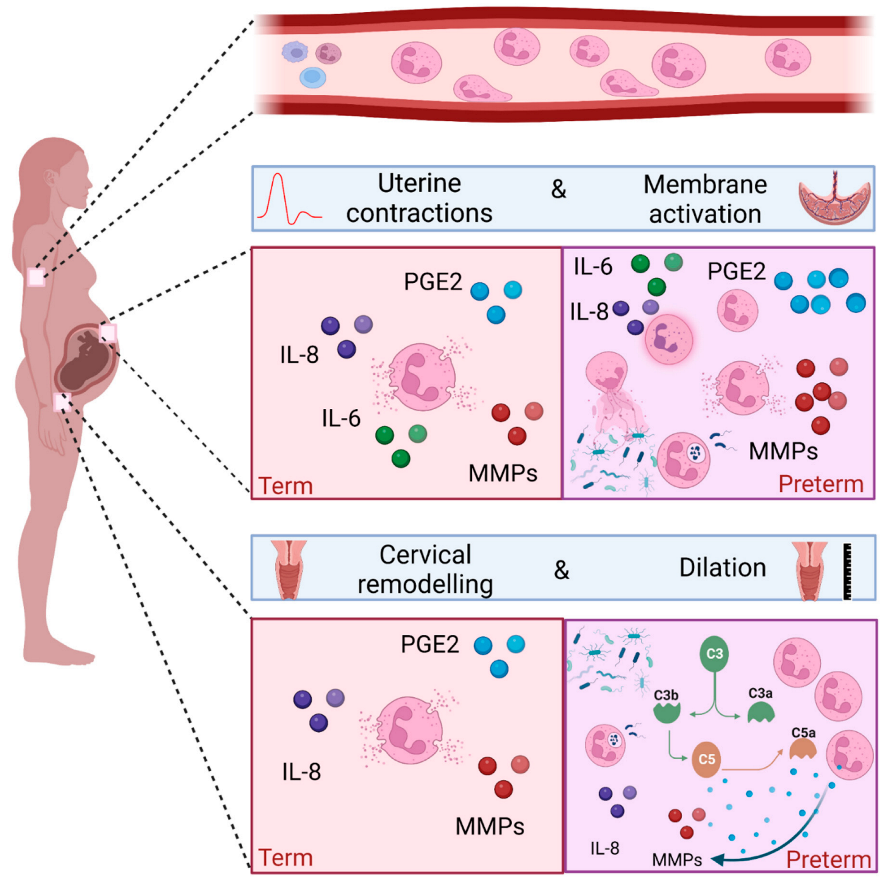
3.1. Peripheral Blood Neutrophils and Term Labour
The onset of labour is consistently associated with neutrophilia [12,64][12][41]. Furthermore, peripheral blood neutrophils taken from women in labour show signs of increased activation compared to women not in established labour [11]. In addition, an increase in markers of migration such as CD11a/b and CD62L is seen in vivo, and in vitro studies have confirmed the increased migratory capacity of neutrophils taken from women in labour [12,64][12][41].
3.2. Neutrophils and the Uterus in Term Labour
Neutrophil infiltration of the myometrium is seen during labour [52,61][29][38] and unsurprisingly this coincides with an increase in cell adhesion molecule expression to aid transmigration [12]. Expression of the chemoattractant CXCL8 mRNA was found to be higher in myometrium of women at term during labour compared to term not in labour, which may explain in part the increased abundance of neutrophils during labour [65][42]. Consistent with this, a parallel increase in IL-8 concentrations and neutrophil counts are seen in the lower uterine segment in women who are in active labour. This is also associated with increased concentrations of matrix metalloproteinases 8 and 9 [66][43].3.3. Neutrophils and the Foetal Membranes in Term Labour
Leukocytes, including neutrophils, are known to infiltrate foetal membranes at the time of term labour [61[38][39],62], and this is accompanied by an increase in the concentrations of pro-inflammatory cytokines such as IL-1β, IL-6, IL-8 [61,62][38][39], and pro-labour mediators such as COX-2 and PGE2 synthases [67,68][44][45].3.4. Neutrophils and Cervical Remodelling in Term Labour
To facilitate vaginal delivery, cervical dilation is one of the processes that occurs during labour. It is a complex process involving softening, effacement and ripening of the cervix. Biochemical changes are required, including a decline in collagen synthesis, an increase in collagenase activity, local immune cell infiltration, and increased concentrations of cytokines and prostaglandins [69][46]. Several studies have demonstrated cervical infiltration of neutrophils from biopsies taken at the time of labour [52,61,66,70][29][38][43][47]. Furthermore, the content of the cervical mucous plug is rich in neutrophils [71][48]. There are conflicting opinions about the importance of cervical neutrophils in causing the cervical changes required for labour, since not all human and murine studies have demonstrated increased neutrophil density in the presence of cervical ripening [52,72][29][49].4. The Role of Neutrophils in Preterm Labour
Preterm birth (PTB) is defined as a birth occurring before 37 completed weeks of pregnancy and can be further classified as extremely preterm (<28 weeks), very preterm (28–32 weeks), and moderate to late preterm (32–36 weeks). There are 15 million babies born preterm each year, with global rates averaging 10%, although significant variations (5–18%) exist depending on geographical location [76][50]. PTB is the biggest cause of childhood mortality under the age of 5, with morbidity and mortality increasing with decreasing gestational age at delivery [77][51]. Roughly two thirds of births are spontaneous, with women presenting either with preterm prelabour rupture of membranes (PPROM), or with uterine contractions and cervical dilation. The causes of preterm birth are multifactorial, but the most common causal factors are infection and/or inflammation. Extreme and very preterm birth are most likely to have evidence of infection and/or inflammation, and babies born with evidence of foetal inflammatory response have a worse prognosis for any given gestational age. Since neutrophils play a major role in infection and inflammation, they are also likely to play a key role in infection and inflammation in the context of preterm birth. The most common source of intrauterine infection is ascending pathogenic microbes from the cervical-vaginal interface [78][52]. The presence of intrauterine infection is associated with neutrophil infiltration and pro-inflammatory cytokine production [79][53]. PPROM often presents with or leads to clinical signs of chorioamnionitis, with neutrophil invasion of the chorioamnion being the hallmark of histological chorioamnionitis [80][54]. Evidence also exists to support the role of neutrophils in driving local inflammation at the cervical-vaginal interface in women who deliver preterm [7,81,82,83][7][55][56][57]. Inflammation in the absence of infection, also referred to as sterile inflammation, has also been widely reported in the context of preterm birth [84,85,86][58][59][60]. DAMPs, also known as alarmins, are endogenous molecules that send a danger message as part of a response to inflammation. Alarmins that have been commonly associated with preterm labour include high mobility group box 1 (HMGB1), IL-1α, and cell free DNA. HMGB1 [87,88][61][62] and IL-1 α [89,90][63][64] concentrations are higher in amniotic fluid of women who have evidence of sterile inflammation and deliver preterm. Cell free DNA activates Toll-like receptor 9 (TLR-9), and animal models support the concept of cell free DNA leading to preterm delivery via leukocyte migration and inflammation at the maternal–foetal interface [91,92][65][66]. Products of the microbiota can be translocated into the circulation, priming and enhancing neutrophil’s function [108][67]. The use of peripheral blood neutrophil concentrations as a predictor of preterm birth has been explored in women presenting in threatened preterm labour and PPROM, using the neutrophil-to-lymphocyte ratio (NLR) and total neutrophil counts [109,110][68][69]. Several studies show an increase in neutrophil counts and the NLR in women who subsequently deliver preterm [12,111][12][70]. Peripheral blood neutrophils exhibit a more activated immunophenotype in women who deliver preterm. Gervasi et al. collected peripheral blood from women who had a healthy pregnancy and subsequent term labour and compared the granulocyte phenotype with women who delivered preterm. Using flow cytometry, they identified that granulocytes expressed higher levels of CD11b, CD15 and CD66 in women who delivered preterm [112][71]. The neutrophils migrate towards a chemotactic gradient, with in vitro evidence demonstrating the chemotactic effect of both unstimulated and LPS stimulated foetal membranes [118][72]. Conditioned media from LPS stimulated foetal membranes leads to the release of cytokines, chemokines, and reactive oxygen species from neutrophils, as well as neutrophil degranulation and NET release [118][72]. No significant differences were seen in neutrophil concentrations between women who delivered at term compared to preterm, however the study was likely to be underpowered for this outcome as only six women delivered preterm. RNA-seq was performed on cervical neutrophils from a subset of the cohort (n = 9). Despite small numbers, the expression of genes involved in neutrophil activation and degranulation negatively correlated with the presence of G. vaginalis and positively correlated with the presence of L. iners in matched vaginal swabs. WeIt haves shown that in women at high risk of preterm birth, neutrophils are more likely to be present at the cervical-vaginal interface if the microbial composition is one of high risk of preterm birth (CST III/L. iners, or CST IV, diverse), compared to low risk (CST I/L. crispatus, CST II/L. gasseri, CST V/L. jensenii) [107][73]. Furthermore, in women that have detectable live cervical neutrophils, there are higher concentrations of pro-inflammatory mediators such as C3b, C1q and C4b in the cervical-vaginal fluid. Taken together, these data suggest a plausible role for cervical neutrophils in microbial driven cervical shortening and PTB [107][73].5. Conclusions
Neutrophils are polymorphonuclear cells and are the most predominant circulating innate immune cells. They play a key role in both inflammation and infection, with effector functions that lead to direct and indirect cell death and microbial clearance. Activated neutrophils secrete pro-inflammatory mediators such as cytokines, proteases and collagenases, and pro-labour mediators such as COX-2 and PGE2. These mediators are required for the physiological processes of healthy term labour; cervical remodelling, uterine contractility, and foetal membrane rupture. However, in cases of microbial driven preterm birth, the premature recruitment of neutrophils into the cervix, uterus and foetal membranes, combined with increased activation, are likely to play a key role in triggering PPROM and PTB.References
- Liew, P.X.; Kubes, P. The Neutrophil’s Role During Health and Disease. Physiol. Rev. 2019, 99, 1223–1248.
- Xie, X.; Shi, Q.; Wu, P.; Zhang, X.; Kambara, H.; Su, J.; Yu, H.; Park, S.-Y.; Guo, R.; Ren, Q.; et al. Single-Cell Transcriptome Profiling Reveals Neutrophil Heterogeneity in Homeostasis and Infection. Nat. Immunol. 2020, 21, 1119–1133.
- Elghetany, M.T. Surface Antigen Changes during Normal Neutrophilic Development: A Critical Review. Blood Cells. Mol. Dis. 2002, 28, 260–274.
- Elghetany, M.T.; Lacombe, F. Physiologic Variations in Granulocytic Surface Antigen Expression: Impact of Age, Gender, Pregnancy, Race, and Stress. J. Leukoc. Biol. 2004, 75, 157–162.
- Lakschevitz, F.S.; Hassanpour, S.; Rubin, A.; Fine, N.; Sun, C.; Glogauer, M. Identification of Neutrophil Surface Marker Changes in Health and Inflammation Using High-Throughput Screening Flow Cytometry. Exp. Cell Res. 2016, 342, 200–209.
- Cloke, T.; Munder, M.; Taylor, G.; Müller, I.; Kropf, P. Characterization of a Novel Population of Low-Density Granulocytes Associated with Disease Severity in HIV-1 Infection. PLoS ONE 2012, 7, e48939.
- Deng, Y.; Ye, J.; Luo, Q.; Huang, Z.; Peng, Y.; Xiong, G.; Guo, Y.; Jiang, H.; Li, J. Low-Density Granulocytes Are Elevated in Mycobacterial Infection and Associated with the Severity of Tuberculosis. PLoS ONE 2016, 11, e0153567.
- Yizengaw, E.; Getahun, M.; Tajebe, F.; Cruz Cervera, E.; Adem, E.; Mesfin, G.; Hailu, A.; Van der Auwera, G.; Yardley, V.; Lemma, M.; et al. Visceral Leishmaniasis Patients Display Altered Composition and Maturity of Neutrophils as Well as Impaired Neutrophil Effector Functions. Front. Immunol. 2016, 7, 517.
- Denny, M.F.; Yalavarthi, S.; Zhao, W.; Thacker, S.G.; Anderson, M.; Sandy, A.R.; McCune, W.J.; Kaplan, M.J. A Distinct Subset of Proinflammatory Neutrophils Isolated from Patients with Systemic Lupus Erythematosus Induces Vascular Damage and Synthesizes Type I Interferons. J. Immunol. 2010, 184, 3284–3297.
- von Dadelszen, P.; Watson, R.W.G.; Noorwali, F.; Marshall, J.C.; Parodo, J.; Farine, D.; Lye, S.J.; Ritchie, J.W.K.; Rotstein, O.D. Maternal Neutrophil Apoptosis in Normal Pregnancy, Preeclampsia, and Normotensive Intrauterine Growth Restriction. Am. J. Obstet. Gynecol. 1999, 181, 408–414.
- Zhang, J.; Shynlova, O.; Sabra, S.; Bang, A.; Briollais, L.; Lye, S.J. Immunophenotyping and Activation Status of Maternal Peripheral Blood Leukocytes during Pregnancy and Labour, Both Term and Preterm. J. Cell. Mol. Med. 2017, 21, 2386–2402.
- Yuan, M.; Jordan, F.; McInnes, I.B.; Harnett, M.M.; Norman, J.E. Leukocytes Are Primed in Peripheral Blood for Activation during Term and Preterm Labour†. Mol. Hum. Reprod. 2009, 15, 713–724.
- Mor, G.; Aldo, P.; Alvero, A.B. The Unique Immunological and Microbial Aspects of Pregnancy. Nat. Rev. Immunol. 2017, 17, 469–482.
- Sykes, L.; MacIntyre, D.A.; Yap, X.J.; Teoh, T.G.; Bennett, P.R. The Th1:Th2 Dichotomy of Pregnancy and Preterm Labour. Mediators Inflamm. 2012, 2012, 967629.
- Racicot, K.; Kwon, J.-Y.; Aldo, P.; Silasi, M.; Mor, G. Understanding the Complexity of the Immune System during Pregnancy. Am. J. Reprod. Immunol. 2014, 72, 107–116.
- Aghaeepour, N.; Ganio, E.A.; Mcilwain, D.; Tsai, A.S.; Tingle, M.; Van Gassen, S.; Gaudilliere, D.K.; Baca, Q.; McNeil, L.; Okada, R.; et al. An Immune Clock of Human Pregnancy. Sci. Immunol. 2017, 2, eaan2946.
- Griffin, J.F.T.; Beck, I. A Longitudinal Study of Leucocyte Numbers and Mitogenesis during the Last Ten Weeks of Human Pregnancy. J. Reprod. Immunol. 1983, 5, 239–247.
- Luppi, P.; Haluszczak, C.; Betters, D.; Richard, C.A.H.; Trucco, M.; DeLoia, J.A. Monocytes Are Progressively Activated in the Circulation of Pregnant Women. J. Leukoc. Biol. 2002, 72, 874–884.
- Chandra, S.; Tripathi, A.K.; Mishra, S.; Amzarul, M.; Vaish, A.K. Physiological Changes in Hematological Parameters During Pregnancy. Indian J. Hematol. Blood Transfus. 2012, 28, 144–146.
- Lampé, R.; Kövér, Á.; Szűcs, S.; Pál, L.; Árnyas, E.; Ádány, R.; Póka, R. Phagocytic Index of Neutrophil Granulocytes and Monocytes in Healthy and Preeclamptic Pregnancy. J. Reprod. Immunol. 2015, 107, 26–30.
- Pramanik, S.S.; Pramanik, T.; Mondal, S.C.; Chanda, R. Number, Maturity and Phagocytic Activity of Neutrophils in the Three Trimesters of Pregnancy. East. Mediterr. Health J. 2007, 13, 862–864.
- Sacks, G.P.; Studena, K.; Sargent, I.L.; Redman, C.W.G. Normal Pregnancy and Preeclampsia Both Produce Inflammatory Changes in Peripheral Blood Leukocytes Akin to Those of Sepsis. Am. J. Obstet. Gynecol. 1998, 179, 80–86.
- Naccasha, N.; Gervasi, M.-T.; Chaiworapongsa, T.; Berman, S.; Yoon, B.H.; Maymon, E.; Romero, R. Phenotypic and Metabolic Characteristics of Monocytes and Granulocytes in Normal Pregnancy and Maternal Infection. Am. J. Obstet. Gynecol. 2001, 185, 1118–1123.
- Köstlin, N.; Kugel, H.; Spring, B.; Leiber, A.; Marmé, A.; Henes, M.; Rieber, N.; Hartl, D.; Poets, C.F.; Gille, C. Granulocytic Myeloid Derived Suppressor Cells Expand in Human Pregnancy and Modulate T-Cell Responses. Eur. J. Immunol. 2014, 44, 2582–2591.
- Farias-Jofre, M.; Romero, R.; Galaz, J.; Xu, Y.; Tao, L.; Demery-Poulos, C.; Arenas-Hernandez, M.; Bhatti, G.; Liu, Z.; Kawahara, N.; et al. Pregnancy Tailors Endotoxin-Induced Monocyte and Neutrophil Responses in the Maternal Circulation. Inflamm. Res. 2022, 71, 653–668.
- Bert, S.; Ward, E.J.; Nadkarni, S. Neutrophils in Pregnancy: New Insights into Innate and Adaptive Immune Regulation. Immunology 2021, 164, 665–676.
- Gomez-Lopez, N.; Romero, R.; Xu, Y.; Miller, D.; Leng, Y.; Panaitescu, B.; Silva, P.; Faro, J.; Alhousseini, A.; Gill, N.; et al. The Immunophenotype of Amniotic Fluid Leukocytes in Normal and Complicated Pregnancies. Am. J. Reprod. Immunol. 2018, 79, e12827.
- Thomson, A.J.; Telfer, J.F.; Young, A.; Campbell, S.; Stewart, C.J.; Cameron, I.T.; Greer, I.A.; Norman, J.E. Leukocytes Infiltrate the Myometrium during Human Parturition: Further Evidence That Labour Is an Inflammatory Process. Hum. Reprod. Oxf. Engl. 1999, 14, 229–236.
- Sakamoto, Y.; Moran, P.; Bulmer, J.N.; Searle, R.F.; Robson, S.C. Macrophages and Not Granulocytes Are Involved in Cervical Ripening. J. Reprod. Immunol. 2005, 66, 161–173.
- Word, R.A.; Li, X.-H.; Hnat, M.; Carrick, K. Dynamics of Cervical Remodeling during Pregnancy and Parturition: Mechanisms and Current Concepts. Semin. Reprod. Med. 2007, 25, 069–079.
- Myers, D.A. The Recruitment and Activation of Leukocytes into the Immune Cervix: Further Support That Cervical Remodeling Involves an Immune and Inflammatory Mechanism. Biol. Reprod. 2012, 87, 107.
- Golightly, E.; Jabbour, H.N.; Norman, J.E. Endocrine Immune Interactions in Human Parturition. Mol. Cell. Endocrinol. 2011, 335, 52–59.
- Liao, J.B.; Buhimschi, C.S.; Norwitz, E.R. Normal Labor: Mechanism and Duration. Obstet. Gynecol. Clin. N. Am. 2005, 32, 145–164.
- Terzidou, V. Biochemical and Endocrinological Preparation for Parturition. Best Pract. Res. Clin. Obstet. Gynaecol. 2007, 21, 729–756.
- Sennström, M.B.; Ekman, G.; Westergren-Thorsson, G.; Malmström, A.; Byström, B.; Endrésen, U.; Mlambo, N.; Norman, M.; Ståbi, B.; Brauner, A. Human Cervical Ripening, an Inflammatory Process Mediated by Cytokines. Mol. Hum. Reprod. 2000, 6, 375–381.
- Challis, J.R.; Lockwood, C.J.; Myatt, L.; Norman, J.E.; Strauss, J.F.; Petraglia, F. Inflammation and Pregnancy. Reprod. Sci. 2009, 16, 206–215.
- Wright, H.L.; Moots, R.J.; Bucknall, R.C.; Edwards, S.W. Neutrophil Function in Inflammation and Inflammatory Diseases. Rheumatology 2010, 49, 1618–1631.
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte Density and Pro-inflammatory Cytokine Expression in Human Fetal Membranes, Decidua, Cervix and Myometrium before and during Labour at Term. Mol. Hum. Reprod. 2003, 9, 41–45.
- Young, A.; Thomson, A.J.; Ledingham, M.; Jordan, F.; Greer, I.A.; Norman, J.E. Immunolocalization of Proinflammatory Cytokines in Myometrium, Cervix, and Fetal Membranes During Human Parturition at Term1. Biol. Reprod. 2002, 66, 445–449.
- Sakamoto, Y.; Moran, P.; Searle, R.F.; Bulmer, J.N.; Robson, S.C. Interleukin-8 Is Involved in Cervical Dilatation but Not in Prelabour Cervical Ripening. Clin. Exp. Immunol. 2004, 138, 151–157.
- Delgado, I.; Neubert, R.; Dudenhausen, J.W. Changes in White Blood Cells during Parturition in Mothers and Newborn. Gynecol. Obstet. Investig 1994, 38, 227–235.
- Bollopragada, S.; Youssef, R.; Jordan, F.; Greer, I.; Norman, J.; Nelson, S. Term Labor Is Associated with a Core Inflammatory Response in Human Fetal Membranes, Myometrium, and Cervix. Am. J. Obstet. Gynecol. 2009, 200, 104.e1–104.e11.
- Winkler, M.; Fischer, D.-C.; Ruck, P.; Marx, T.; Kaiserling, E.; Oberpichler, A.; Tschesche, H.; Rath, W. Parturition at Term: Parallel Increases in Interleukin-8 and Proteinase Concentrations and Neutrophil Count in the Lower Uterine Segment. Hum. Reprod. 1999, 14, 1096–1100.
- Osman, I.; Young, A.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte Density and Proinflammatory Mediator Expression in Regional Human Fetal Membranes and Decidua Before and During Labot at Term. J. Soc. Gynecol. Investig. 2006, 13, 97–103.
- Takahashi, N.; Okuno, T.; Fujii, H.; Makino, S.; Takahashi, M.; Ohba, M.; Saeki, K.; Itakura, A.; Takeda, S.; Yokomizo, T. Up-Regulation of Cytosolic Prostaglandin E Synthase in Fetal-Membrane and Amniotic Prostaglandin E2 Accumulation in Labor. PLoS ONE 2021, 16, e0250638.
- Norman, J.E. Cervical Function and Prematurity. Best Pract. Res. Clin. Obstet. Gynaecol. 2007, 21, 791–806.
- Bokström, H.; Brännström, M.; Alexandersson, M.; Norström, A. Leukocyte Subpopulations in the Human Uterine Cervical Stroma at Early and Term Pregnancy. Hum. Reprod. 1997, 12, 586–590.
- Hein, M.; Petersen, A.C.; Helmig, R.B.; Uldbjerg, N.; Reinholdt, J. Immunoglobulin Levels and Phagocytes in the Cervical Mucus Plug at Term of Pregnancy. Acta Obstet. Gynecol. Scand. 2005, 84, 734–742.
- Timmons, B.C.; Mahendroo, M.S. Timing of Neutrophil Activation and Expression of Proinflammatory Markers Do Not Support a Role for Neutrophils in Cervical Ripening in the Mouse1. Biol. Reprod. 2006, 74, 236–245.
- Blencowe, H.; Cousens, S.; Oestergaard, M.Z.; Chou, D.; Moller, A.-B.; Narwal, R.; Adler, A.; Vera Garcia, C.; Rohde, S.; Say, L.; et al. National, Regional, and Worldwide Estimates of Preterm Birth Rates in the Year 2010 with Time Trends since 1990 for Selected Countries: A Systematic Analysis and Implications. Lancet 2012, 379, 2162–2172.
- Liu, L.; Oza, S.; Hogan, D.; Perin, J.; Rudan, I.; Lawn, J.E.; Cousens, S.; Mathers, C.; Black, R.E. Global, Regional, and National Causes of Child Mortality in 2000–13, with Projections to Inform Post-2015 Priorities: An Updated Systematic Analysis. Lancet 2015, 385, 430–440.
- Agrawal, V.; Hirsch, E. Intrauterine Infection and Preterm Labor. Semin. Fetal. Neonatal Med. 2012, 17, 12–19.
- Kim, C.J.; Romero, R.; Chaemsaithong, P.; Chaiyasit, N.; Yoon, B.H.; Kim, Y.M. Acute Chorioamnionitis and Funisitis: Definition, Pathologic Features, and Clinical Significance. Am. J. Obstet. Gynecol. 2015, 213, S29–S52.
- Steel, J.H.; O’Donoghue, K.; Kennea, N.L.; Sullivan, M.H.F.; Edwards, A.D. Maternal Origin of Inflammatory Leukocytes in Preterm Fetal Membranes, Shown by Fluorescence in Situ Hybridisation. Placenta 2005, 26, 672–677.
- Yamada, T.; Minakami, H.; Matsubara, S.; Yatsuda, T.; Sato, I. Changes in Polymorphonuclear Leukocytes in the Vagina of Patients with Preterm Labor. Gynecol. Obstet. Investig 1998, 45, 32–34.
- Hezelgrave, N.L.; Seed, P.T.; Chin-Smith, E.C.; Ridout, A.E.; Shennan, A.H.; Tribe, R.M. Cervicovaginal Natural Antimicrobial Expression in Pregnancy and Association with Spontaneous Preterm Birth. Sci. Rep. 2020, 10, 12018.
- Whitworth, M.K.; Pafilis, I.; Vince, G.; Quenby, S. Cervical Leukocyte Sub-Populations in Idiopathic Preterm Labour. J. Reprod. Immunol. 2007, 75, 48–55.
- Stranik, J.; Kacerovsky, M.; Andrys, C.; Soucek, O.; Bolehovska, R.; Holeckova, M.; Matulova, J.; Jacobsson, B.; Musilova, I. Intra-Amniotic Infection and Sterile Intra-Amniotic Inflammation Are Associated with Elevated Concentrations of Cervical Fluid Interleukin-6 in Women with Spontaneous Preterm Labor with Intact Membranes. J. Matern. Fetal Neonatal Med. 2021, 1–9.
- Gomez-Lopez, N.; Romero, R.; Varrey, A.; Leng, Y.; Miller, D.; Done, B.; Xu, Y.; Bhatti, G.; Motomura, K.; Gershater, M.; et al. RNA Sequencing Reveals Diverse Functions of Amniotic Fluid Neutrophils and Monocytes/Macrophages in Intra-Amniotic Infection. J. Innate Immun. 2021, 13, 63–82.
- Nadeau-Vallée, M.; Obari, D.; Palacios, J.; Brien, M.-È.; Duval, C.; Chemtob, S.; Girard, S. Sterile Inflammation and Pregnancy Complications: A Review. Reproduction 2016, 152, R277–R292.
- Romero, R.; Miranda, J.; Chaiworapongsa, T.; Korzeniewski, S.J.; Chaemsaithong, P.; Gotsch, F.; Dong, Z.; Ahmed, A.I.; Yoon, B.H.; Hassan, S.S.; et al. Prevalence and Clinical Significance of Sterile Intra-Amniotic Inflammation in Patients with Preterm Labor and Intact Membranes. Am. J. Reprod. Immunol. 2014, 72, 458–474.
- Baumbusch, M.A.; Buhimschi, C.S.; Oliver, E.A.; Zhao, G.; Thung, S.; Rood, K.; Buhimschi, I.A. High Mobility Group-Box 1 (HMGB1) Levels Are Increased in Amniotic Fluid of Women with Intra-Amniotic Inflammation-Determined Preterm Birth, and the Source May Be the Damaged Fetal Membranes. Cytokine 2016, 81, 82–87.
- Figueroa, R.; Garry, D.; Elimian, A.; Patel, K.; Sehgal, P.B.; Tejani, N. Evaluation of Amniotic Fluid Cytokines in Preterm Labor and Intact Membranes. J. Matern. Fetal Neonatal Med. 2005, 18, 241–247.
- Romero, R.; Brody, D.T.; Oyarzun, E.; Mazor, M.; King Wu, Y.; Hobbins, J.C.; Durum, S.K. Infection and Labor: III. Interleukin-1: A Signal for the Onset of Parturition. Am. J. Obstet. Gynecol. 1989, 160, 1117–1123.
- Thaxton, J.E.; Romero, R.; Sharma, S. TLR9 Activation Coupled to IL-10 Deficiency Induces Adverse Pregnancy Outcomes. J. Immunol. 2009, 183, 1144–1154.
- Scharfe-Nugent, A.; Corr, S.C.; Carpenter, S.B.; Keogh, L.; Doyle, B.; Martin, C.; Fitzgerald, K.A.; Daly, S.; O’Leary, J.J.; O’Neill, L.A.J. TLR9 Provokes Inflammation in Response to Fetal DNA: Mechanism for Fetal Loss in Preterm Birth and Preeclampsia. J. Immunol. 2012, 188, 5706–5712.
- Clarke, T.B.; Davis, K.M.; Lysenko, E.S.; Zhou, A.Y.; Yu, Y.; Weiser, J.N. Recognition of Peptidoglycan from the Microbiota by Nod1 Enhances Systemic Innate Immunity. Nat. Med. 2010, 16, 228–231.
- Ozel, A.; Alici Davutoglu, E.; Yurtkal, A.; Madazli, R. How Do Platelet-to-Lymphocyte Ratio and Neutrophil-to-Lymphocyte Ratio Change in Women with Preterm Premature Rupture of Membranes, and Threaten Preterm Labour? J. Obstet. Gynaecol. 2020, 40, 195–199.
- Lakshmi, M.P.A.S.; Sravani, V.L. Role of Neutrophil-Lymphocyte Ratio in Determining the Outcomes of Preterm Premature Rupture of Membranes. Int. J. Reprod. Contracept. Obstet. Gynecol. 2021, 10, 1617–1620.
- Vakili, S.; Torabinavid, P.; Tabrizi, R.; Shojazadeh, A.; Asadi, N.; Hessami, K. The Association of Inflammatory Biomarker of Neutrophil-to-Lymphocyte Ratio with Spontaneous Preterm Delivery: A Systematic Review and Meta-Analysis. Mediators Inflamm. 2021, 2021, 6668381.
- Gervasi, M.-T.; Chaiworapongsa, T.; Naccasha, N.; Blackwell, S.; Yoon, B.H.; Maymon, E.; Romero, R. Phenotypic and Metabolic Characteristics of Maternal Monocytes and Granulocytes in Preterm Labor with Intact Membranes. Am. J. Obstet. Gynecol. 2001, 185, 1124–1129.
- Tong, M.; Potter, J.A.; Mor, G.; Abrahams, V.M. Lipopolysaccharide-Stimulated Human Fetal Membranes Induce Neutrophil Activation and Release of Vital Neutrophil Extracellular Traps. J. Immunol. 2019, 203, 500–510.
- Molina, B.; Bayar, E.; Lee, Y.S.; Muller, I.; Botto, M.; MacIntyre, D.; Bennett, P.R.; Kropf, P.; Sykes, L. Cervicovaginal Inflammation and Neutrophils Infiltration/Activation in Women at High-Risk of Prematurity. BJOG Int. J. Obstet. Gynaecol. 2022, 129, 47–62.