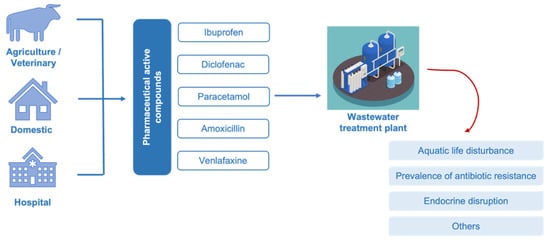
The worldwide access to pharmaceuticals and their continuous release into the environment have raised a serious global concern. Pharmaceuticals remain active even at low concentrations, therefore their occurrence in waterbodies may lead to successive deterioration of water quality with adverse impacts on the ecosystem and human health. To address this challenge, there is an evolving trend toward the search for effective methods to ensure efficient purification of both drinking water and wastewater. Biocatalytic transformation of pharmaceuticals using oxidoreductase enzymes, such as peroxidase and laccase, is a promising environmentally friendly solution for water treatment, where fungal species have been used as preferred producers due to their ligninolytic enzymatic systems.
- oxidoreductases
- pharmaceuticals
- enzyme immobilization
- biodegradation
- wastewater
1. Occurrence and Toxicity of Pharmaceutical Aactive Ccompounds
2. Enzymatic Biodegradation
Enzymes are catalysts that conduct, within the mild conditions of temperature and pH, chemical reactions at a remarkably high rate, efficiency and specificity [15][22]. Several enzyme systems have been used for the efficient transformation and degradation of organic pollutants and have been shown to oxidize and degrade the pollutants into smaller intermediates [16][35]. The literature survey shows that most enzymatic remediation studies use oxidoreductase enzymes [17][18][36,37]. These enzymes are largely produced by the white-rot fungi (WRF), as extracellular enzymes for the degradation of lignin. WRF and their ligninolytic enzymes, namely lignin peroxidase (LiP), manganese peroxidase (MnP), versatile peroxidase (VP) and Lac, have been demonstrated to be capable of transforming a wide range of compounds. This ability is a result of the structural similarities of several micropollutants to lignin, as well as the fact that ligninolytic enzymes are substrate-nonspecific [19][38]. Other enzymes are, as well, used as biocatalysts for the remediation of toxic compounds, such as tyrosinase, horseradish peroxidase (HRP) and phenoloxidase [17][36]. Peroxidases (EC 1.11.1.X) are a vast group of heme-containing oxidoreductases, which use hydrogen peroxide as an electron-acceptor to catalyze oxidative reactions [20][39], whereas Lac (EC 1.10.3.2) are glycosylated multi-copper oxidases that catalyzes one-electron oxidation of various substrates associated with the reduction of molecular oxygen to water, via a radical-catalyzed reaction mechanism [21][40]. In contrast to peroxidases that require the supply of hydrogen peroxide, Lac only requests oxygen as the final electron acceptor for the oxidation reaction to occur, offering an alternative green approach for the biodegradation of several PhAC [22][41]. Lac is selective towards phenolic compounds, but due to the non-specificity, they are also able to degrade aromatic amines and related substances, thiol groups, diamines, N-heterocycles and phenothiazines [23][42]. The active sites of Lac enzyme, as represented in Figure 2, contain four copper ions: one type 1 (T1) copper ion and a trinuclear copper cluster (TNC) composed of one type 2 (T2) copper ion coupled to binuclear type 3 (T3) copper ions [24][43]. The T1 copper site is the primary electron-acceptor for electrons offered by the substrate. The electrons are then moved to the TNC through highly conserved His-Cys-His tripeptide. After that, oxygen reacts with the fully reduced enzyme to form a peroxy-intermediate (PI), and then the PI is transformed back into a native intermediate, through a two-electron reduction. Once the fully reduced state is recovered, the final products are released from the TNC site. This results in the production of four radicals by the oxidation of four molecules of substrate, while one molecule of oxygen is reduced to two molecules of water and four protons (H+) are consumed from the solution [25][26][44,45].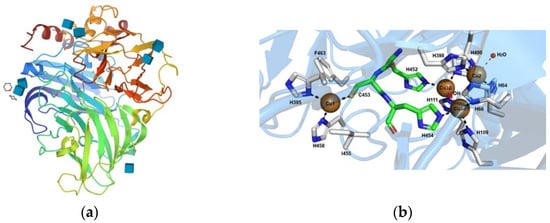
| Compound | Enzyme | Source | PhAC (mg/L) | Reaction Conditions | Enzyme Load (U/L) | Efficiency (%) | Ref. | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diclofenac | LiP | Phanerochaete chrysosporium | 5 | pH 4, 24 mg/L H2O2, 25 °C, 2 h. | 180 | 100 | [34][53] | |||||
| Lac | Trametes versicolor | 1 | pH 6.5, 25 °C, 5 h. | 500 | >90 | [35][54] | ||||||
| Lac | Pycnoporus sanguineus | 100 | pH 5, 25 °C, 8 h. | 100 | 50 | [36][55] | ||||||
| 5,7-Diiodo-8-hydroxyquinoline | Lac | Pycnoporus sanguineus | 100 | pH 5, 25 °C, 3.5 h. | 100 | 78 | [36][55] | |||||
| Carbamazepine | Lac | Trametes versicolor | 1 | pH 6, 35 °C, 24 h. | 60 | 30 | [32][51][37][56] | |||||
| Salicylic acid | Lac | Trametes pubescens | 0.001 | pH 6.9, 25 °C, 24 h. | 100 | >90 | [ | |||||
| Ketoconazole | Trametes versicolor | 38 | 1 mM of HBT | 300 mg/L PhAC. pH 4.5, 45 °C, 6 h. | 1000 | ] | [ | 9857] | ||||
| [ | 67 | ] | [ | 83 | ] | 17-α-ethynyl estradiol | Lac | Trametes pubescens | 0.001 | pH 6.9, 25 °C, 24 h. | 100 | >90 |
| Naproxen | Pleurotus ostreatus | 1 mM of ABTS | [ | 38 | ] | [ | 57] | |||||
| 0.5 mg/L PhAC. 25 °C, 8 h. | 0.26 | 80 | [ | 66 | ] | [82] | Sulfamethoxazole | Lac | Phanerochaete chrysosporium | 10 | pH 4.5, 30 °C, 48 h. | 6076 |
| Olsalazine | Aspergillus aculeatus | 2 mM of ABTS 1, p-Coumaric acid 2 and HBT | 50 | 3 | 100 mg/L PhAC. pH 5, 29 °C, 48 h. | - | 99 1 98 2 94 3 |
[68][39][58] | ||||
| [ | 84 | ] | Lac | Pycnoporus sanguineus | 10 | |||||||
| Atenolol | Trametes versicolor | 0.5 mM of TEMPO | 30 °C, 72 h. | 170 | 29 | 2.7 mg/L PhAC. pH 7, 25 °C, 4 h. | 5000 | 80 | [69][85] |
2.2. Transformation Mechanisms and Toxicity Evaluation
Enzymes transform complex compounds into simpler substances and it is unknown whether pharmaceuticals are metabolized by remediating enzymes to less or more toxic products. There is also a lack of knowledge on the structure of the metabolites resulting from pharmaceuticals degradation process. Therefore, some authors have studied the transformation pathways, as well as metabolites toxicity or estrogenic activity. Considering molecular weight and chemical structure, Kózka et al. [71][87] observed three main types of transformations of a series of antidepressants and immunosuppressants carried out by fungal ligninolytic enzymes. The first one is chemical oxidation and occurred for clomipramine, mianserin, sertraline, fluoxetine and citalopram. The second transformation is straight demethylation or demethylation coupled with other reactions such as oxidation or deamination, and was observed for clomipramine, mianserin, sertraline and venlafaxine transformation. The third type of transformation is the oxidative cleavage of the molecule into two parts of comparable size and was observed for fluoxetine and paroxetine. Kasonga et al. [72][88] proposed the metabolic pathways for ibuprofen (IBP) and CBZ based on detected intermediates by a fungal consortium of Lac, LiP and MnP. The IBP transformation pathway appeared to result from hydroxylation with addition of hydroxyl group to 1,2-dihydroxy-IBP or carboxylation reaction leading to the substitution of the methyl group by carboxylic group to form IBP carboxylic acid. Furthermore, the CBZ metabolic pathway was presented in four routes. The first route proposed was oxidation or hydrolysis to iminostilbene. The second route consisted of oxidation reactions of the carbons on the aromatic benzene group to CBZ-2,3-quinone. The third route combined hydroxylation, hydrolysis and then oxidization to iminoquinone. The fourth and principal metabolic route started with oxidation or epoxidation and the end products were acridone, 10,11-dihydro-10-hydroxy-CBZ and 9-hydroxymethyl-10-carbamoyl acridan. Likewise, according to Naghdi et al. [37][56], 10,11-dihydro-10,11-dihydroxy-CBZ and 10,11-dihydro-10,11-epoxy-CBZ are considered to be the primary metabolites from CBZ oxidation by Lac-ABTS. Moreover, toxicity tests revealed that these products had no estrogenic effect. In another study related to the degradation of DCF by Lac, the authors identified 3′-hydroxydiclofenac, 4′-hydroxydiclofenac, and 5-hydroxydiclofenac as the major transformation products [35][54]. Yang et al. [73][89] demonstrated that immobilized C. unicolor Lac was effective in detoxification of TC antibiotics and identified three transformation products with LC-TOF-MS. According to the authors, TC is first oxidized by Lac to the corresponding ketone and then the amino group is bi-demethylated to form the second transformation product. The final product results from oxidation, followed by water elimination and dehydrogenation. Lac oxidation of SMX in presence of ABTS possibly results in two products from pharmaceutical degradation and a third one from ABTS oxidation by Lac, designated 3-ethyl-6-sulfonate benzothiazolinone imine [64][81]. In another study, Tian et al. [22][41] demonstrated that TC was first oxidized by Lac-ABTS to OTC as the major transformation product and proposed a degradation pathway including deamination, demethylation and dehydration. The assessment of the overall toxicity of the treated effluents is essential to provide a more complete outlook of the environmental relevance of the enzymatic treatments in real practical applications. Several metabolites result from enzymatic water treatment and additional toxicological information by means of bioassays is imperative. These studies are able to describe the ecotoxicity and the estrogenic activity of the resulting metabolites. Presently, there are examples of pharmaceutical enzymatic degradation that produce fewer toxic compounds and lack of estrogenicity effect. A micro-toxicity study with Pseudokirchneriella subcapitata, Candida albicans, Cryptococcus neoformans and Saccharomyces cerevisiae revealed that Lac-HBT treated ketoconazole and its isolated metabolites, such as 1-(4-{4-[2-(2,4-dichloro-phenyl)-2-imidazol-1-ylmethyl-[1,3]dioxolan-4-ylmethoxy]-phenyl}-4-oxy-piperazin-1-yl)-ethanone, suffered a decrease in the toxicity levels. The presence of the oxygen atom in the structure of metabolites reduces their lipophilicity and decreases their toxicity [67][83]. Furthermore, Lac-SA mediated system led to the transformation of CAP in chloramphenicol aldehyde and had less toxicity for microbial growth than mediators vanillin, ABTS and α-naphthol [32][51]. Spina et al. [74][90] showed that crude Lac from Trametes pubescens MUT 2400 was very active against all target micropollutants as ketoprofen, present in real municipal wastewater. Estrogenic analysis and toxicological tests with Raphidocelis subcapitata and Lepidium sativum showed a clear ecotoxicity reduction of treated wastewater. Similarly, Sun et al. [31][50] reported that a concentrated Lac isolated from Trametes hirsuta was capable of effectively metabolize 17b-estradiol (E2) more than 99% and potentially lead to a reduction of the estrogenic activity of E2. Through the combination of 13C-isotope labelling with high-resolution mass spectrometry, the dimers, trimers and tetramers were recognized as the primary by-products of E2 metabolism. Contrarily, Becker et al. [75][91] verified that Lac mediated with SA effectively removes (>50%) a broad range of antibiotics after 24 h. However, this enhanced degradation induces unspecific toxicity. Furthermore, Feng et al. [69][85] verified that the transformation of ATL via Lac/TEMPO-catalyzed reaction greatly reduced the mortalities of zebrafish (Danio rerio) eggs, but the degradation products and the residual TEMPO still possess toxicity (approximately 40%). This transformation mainly involved hydroxylation, carbonylation, C–O bond cleavage and coupling reactions.2.3. Immobilized Biocatalytic System
The practical application of freely suspended enzymes exhibits high activity in the biotransformation processes. Nevertheless, the free form is limited by the low stability and high cost of production for large scale implementation due to the impossibility of recovery [76][77][92,93]. To overcome such limitations, several studies have already demonstrated that enzyme activity and stability can be improved by immobilization on solid supports [17][36]. The stabilization of the peptide structure of the biocatalyst, creating interactions between the enzyme and an immobilization matrix, leads to enzyme stability and resistance improvement towards extreme operational conditions, including strong pH, high temperature or the presence of organic solvents [78][94]. Furthermore, immobilization allows the easy recovery of the enzyme and offers high reusability in several catalytic cycles without significant loss of its unique properties, which reduces operating costs [79][95]. The immobilization strategies are divided into methods based on physical or chemical interactions between enzymes and supports [80][96]. Physical immobilization involves the creation of non-specific interactions via hydrogen bonds, ionic and hydrophobic interactions. The physical methods include entrapment, encapsulation and adsorption, and there is no requirement for the functionalization of the support [17][36]. On the other hand, chemical immobilization includes enzyme attachment to the matrix by covalent binding or cross-linking [81][97]. An example of a covalent binding agent is glutaraldehyde that is capable to react with the amine groups at the surface of both enzymes and support through the formation of Schiff’s bases and Michael’s adducts [82][98]. Moreover, enzymes can be cross-linked to each other or create a cross-linked enzyme aggregate (CLEA) [83][99]. Physical adsorption is simpler and leads to higher final enzyme activity. However, desorption or leakage of the immobilized enzyme is common with cycles of use due to the relatively weak binding forces. Oppositely, chemical immobilization leads to partial deformations in the enzyme molecular shape but offers a robust attachment of enzymes to the support [78][84][94,100]. The characteristics of the support are essential to define the success of the final biocatalyst, therefore the ideal support for usage of industrial applications should be inert, rigid, inexpensive, eco-friendly and present thermal and mechanical resistances [85][24]. The support matrices can be classified according to their chemical composition as inorganic materials, organic materials, hybrids and composite materials. A large diversity of support has been developed due to the search for better stability and scale-up performance. Several recent methods of enzyme immobilization are summarized in Table 4.| Support Material | Enzyme | Immobilization Method | PhAC | Reaction Conditions | Efficiency (%) | Ref. | |
|---|---|---|---|---|---|---|---|
| Poly(l-lactic acid)-co-poly(ε-caprolactone) nanofibers | Lac from Trametes versicolor | Encapsulation | Naproxen | 1 mg/L PhAC. pH 5, 25 °C, 24 h, 100 rpm. |
90 | [77][93] | |
| Poly(l-lactic acid)-co-poly(ε-caprolactone) nanofibers | Lac from Trametes versicolor | Encapsulation | Diclofenac | 1 mg/L PhAC. pH 3, 25 °C, 24 h, 100 rpm. |
90 | [77][93] | |
| Polyvinylidene fluoride membrane with multi-walled carbon nanotubes | Lac from Trametes hirsuta | Covalent bonding | 5 mg/L PhAC. pH 5, 25 °C, 4 h. |
95 | [78][94] | ||
| Titania nanoparticles | Lac from Pycnoporus sanguineus CS43 | Covalent bonding | 10 mg/L PhAC. pH 4, 25 °C, 4 h. |
50 | [86][101] | ||
| Micro-biochar from pine wood (PW) and pig manure (PM) | Lac from Trametes versicolor | Covalent bonding | 0.5 mg/L PhAC. pH 6.5, 25 °C, 5 h (PW) or 2 h (PM). |
99 | [82][98] | ||
| Chitosan macro-beads | Lac from Trametes versicolor | Covalent bonding | 50 mg/L PhAC. pH 3, 25 °C, 4 h, 1:1 M ratio for ABTS:drug. |
90 | [30][49] | ||
| CLEA | Lac from Trametes versicolor | Cross-linking | 0.001 mg/L PhAC. pH 5, 24 h, 22 °C. |
90 | [83][99] | ||
| Polyacrylonitrile−biochar composite nanofibrous membrane |
Lac from Trametes versicolor | Covalent bonding | 0.2 mg/L PhAC, pH 4, 8 h, 35 °C. |
73 | [87][102] | ||
| Polyacrylonitrile−biochar composite nanofibrous membrane |
Lac from Trametes versicolor | Covalent bonding | Chlortetracycline | 0.2 mg/L PhAC, pH 4, 8 h, 35 °C. | [ | 40][59] | |
| 17-β-estradiol | Lac | Trametes hirsuta | 5 | pH 5, 25 °C, 120 min. | 5000 | 99 | [31][50] |
| Lac | Trametes pubescens | 0.001 | pH 6.9, 25 °C, 24 h. | 100 | >90 | [38][57] | |
| Tetracycline 1 Oxytetracycline 2 |
MnP | Phanerochaete chrysosporium | 50 | pH 4.8, 0.1 mM Mn2+, 0.1 mM H2O2, 37 °C, 4 h. | 40 | 73 1 84 2 |
[41][60] |
| Acetaminophen | Lac | Bjerkandera adusta TBB-03 | 20 | pH 5–7, 25 °C, 2 h. | 270 | 100 | [42][61] |
| HRP | Horseradish | 6 | pH 7.4, 400 μM H2O2, 25 °C, 4 h. | 12800 | 100 | [2][8] | |
| Triclosan | Lac | Trametes versicolor | 3 | pH 6, 25 °C, 4 h. | 2000 | 52 | [43][62] |
| Doxorubicin | Lac | Trametes Versicolor | 0.25 | pH 7, 30 °C, 2 h. | 900 | 100 | [44][21] |
| Imipramine | Lac | Paraconiothyrium variabile | 0.12 | pH 5, 37 °C, 6 h. | 1600 | 98 | [45][20] |
2.1. Laccase-Mediator Catalyzed System
Lac possess a relatively low redox potential (≤0.8 V) compared to peroxidase (>1 V). The redox potential difference between the substrate and the enzyme T1 copper affects the oxidative capacity of Lac. For example, Lac efficiently promotes single-electron oxidation of phenols. Accordingly, non-phenolic compounds, with redox potential above 1.3 V, are not directly prone to oxidation by Lac [62][79]. This lack of affinity of Lac is generally influenced by the distribution of functional groups in the chemical structure of the substrates. Compounds with electron withdrawing groups (EWG) such as carboxylic (-COOH), amide (-CONR2), halogen (-X) and nitro (-NO2) have lower enzyme affinity due to the less susceptible to oxidative catabolism [63][80]. The degradation efficiency of pollutants using Lac can be enhanced by the addition of redox mediators that are easily oxidized by the enzyme to free radicals. The presence of these small molecular weight compounds allows Lac to overcome a kinetic barrier and increase the spectrum of pollutants potentially degraded, as the mediator species have higher redox potentials. These compounds act as an “electron shuttle”, enabling the oxidation of complex substrates by highly reactive radicals that result from mediator oxidation by Lac. These radicals may return to their parent compound through reduction during the oxidation of the target pollutant [33][64][52,81]. Table 2 presents a list of redox mediators and their related information. The hydrogen atom transfer (HAT), electron transfer (ET) and ionic mechanisms are the primary mechanisms for mediator oxidation of a compound. The oxidation mechanism of the 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt (ABTS) and 2, 2, 6, 6-tetramethyl-1-piperidinyloxy (TEMPO) radicals follows ET and ionic mechanisms, respectively. As well as p-coumaric acid, hydroxybenzotriazole (HBT), N-hydroxyphthalimide (HPI) and syringaldehyde (SA) follow HAT oxidation mechanism [65][18].| Oxidation Mechanism | Redox Mediator | Origin | Type of Mediator | Free Radical Generated |
|---|---|---|---|---|
| Electron transfer | 2,2′-azino-bis (3-ethylbenzothiazoline-6-sulfonic acid) diammonium salt | Synthetic | ABTS | ABTS |
| Synthetic | ||||
| N-O | ||||
| N=O | ||||
| • | ||||
| Oxo-ammonium |
| Compound | Lac Source | Mediator | Reaction Conditions | Enzyme Load (U/L) | Efficiency (%) | Ref. | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Diclofenac | Trametes versicolor | 1 mM of HBT 1 and SA 2 | 0.1 mg/L PhAC. 25 °C, 24 h. | •+ ABTS2+ |
||||||
| 1440 | >95 | 1 | 80 | 2 | [33][52] | Hydrogen atom transfer | Hydroxybenzotriazole | Synthetic | N–OH | |
| Carbamazepine | Trametes versicolor | 0.018 mM of ABTS | 1 mg/L PhAC. pH 6, 35 °C, 24 h. | =N–O• Aminoxyl |
||||||
| 60 | 95 | [ | 37 | ] | [ | 56] | N-hydroxyphthalimide | Synthetic | N–OH | |
| Sulfamethoxazole | Trametes versicolor | =N–O | • | 0.5 mM ABTS, SA and AS Aminoxyl |
||||||
| 20-25 mg/L PhAC. pH 6–7, 25 °C, 2–6 h. | 560 | 100 | [ | 64 | ] | [81] | Violuric acid | Natural | N–OH | =N–O |
| Tetracycline Oxytetracycline |
Pycnoporus sp. SYBC-L10 | • | Aminoxyl |
|||||||
| 1 mM of ABTS | 50 mg/L PhAC. pH 6, 5 min, 0 °C. | 10,000 | 100 | [ | 22 | ][41] | Vanillin | Natural | C6H4(OH)(OCH3) | C6H5O• Phenoxyl |
| Syringaldehyde | Natural | C6 | ||||||||
| Chloramphenicol | Trametes hirsuta | 0.5 mM of SA, vanillin, ABTS and α-naphthol | 10 mg/L PhAC. pH 5, 25 °C, 48 h. | 50,220 | 100H4(OH)(OCH3) | C6H5O• Phenoxyl |
||||
| Acetosyringone | Natural | C6H4(OH)(OCH3) | C6H5O• Phenoxyl |
|||||||
| p-coumaric acid | Natural | C6H4(OH)(OCH3) | C6H5O• Phenoxyl |
|||||||
| Ionic oxidation | 2,2,6,6-tetramethylpiperidinyloxyl | 63 | [ | 87 | ] | [102] | ||||
| Polyvinylidene fluoride membrane with multi-walled carbon nanotubes | Lac from Trametes hirsuta | Covalent bonding | Carbamazepine | 5 mg/L PhAC. pH 5, 25 °C, 48 h. |
27 | [78][94] | ||||
| Magnetite nanoparticles |
HRP LiP |
Adsorption | 0.35 mg/L PhAC. pH 3, 55 °C, 3 days. |
100 | [88][103] | |||||
| Polyimide aerogels | Lac from Trametes versicolor | Covalent bonding | 0.02 mg/L PhAC. pH 3, 25 °C, 24 h, 200 rpm. |
74 | [89][104] | |||||
| Pinewood nanobiochar | Lac from Trametes versicolor | Adsorption | 0.02 mg/L PhAC. pH 3.5, 25 °C, 24 h, 200 rpm. |
80 | [76][92] | |||||
| Polyamide/polyethylenimine nanofibers | Lac from Trametes versicolor | Covalent bonding | Triclosan | 10 mg/L PhAC. pH 7, 25 °C, 20 h, 80 rpm. |
74 | [90][105] | ||||
| Titania nanoparticles | Lac from Pycnoporus sanguineus CS43 | Covalent bonding | Acetaminophen | 10 mg/L PhAC. pH 4, 25 °C, 4 h. |
90 | [86][101] | ||||
| Commercial silica gel particles | Lac from Trametes versicolor | Covalent bonding | Sulfamethoxazole | 20 mg/L PhAC. pH 7, 25 °C, 0.5 h. 520 μM of ABTS. | 53 | [91][106] | ||||
| Commercial silica gel particles | Lac from Trametes versicolor | Covalent bonding | Amoxicillin | 20 mg/L PhAC. pH 7, 25 °C, 4 h. 520 μM of ABTS. | 80 | [91][106] | ||||
| M-CLEA | Lac from Cerrena unicolor | Cross-linking | Tetracycline | 100 mg/L PhAC. pH 6, 25 °C, 48 h. |
100 | [73][89] | ||||
| Mesostructured cellular foam | Lac from Trametes versicolor | Adsorption | 1 mg/L PhAC. pH 5, 25 °C, 1 h. |
100 | [92][107] | |||||
| Bentonite-derived mesoporous materials | Lac from Trametes versicolor | Adsorption | 10 mg/L PhAC. 30 °C, 3 h. | 60 | [84][100] | |||||
| Cellulose beads |
Lac from Trametes versicolor | Covalent bonding | Indole | 15 mg/L PhAC. pH 5, 30 °C, 18 h. |
100 | [93][108] | ||||
| Polypropylene beads | Lac from Myceliophthora thermophila | Adsorption | Morphine | 1 mg/L PhAC. pH 6, 25 °C, 0.5 h. |
100 | [94][109] | ||||
| Graphene oxide and alginate matrix | Lac from Aspergillus niger | Adsorption/entrapment | Cetirizine dihydrochloride | 20 mg/L PhAC. pH 4.5, 25 °C, 1 h. |
100 | [95][23] | ||||
| Pristine few layers graphene | Lac from Trametes versicolor | Adsorption | Labetalol hydrochloride | 1 mg/L PhAC pH 7, 25 °C, 1.5 h. 5 μM of ABTS. | 100 | [96][110] |