Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Zhi Chen and Version 2 by Amina Yu.
Beta-oxidation(β-oxidation) is an important metabolic process involving multiple steps by which fatty acid molecules are broken down to produce energy. The very long-chain fatty acids (VLCFAs), a type of fatty acid (FA), are usually highly toxic when free in vivo, and their oxidative metabolism depends on the peroxisomal β-oxidation. Although peroxisomal β-Oxidation attracts less research than mitochondria, the importance of the peroxisomal β-oxidation molecular mechanism can still be spotted from some mechanisms involved in upstream regulation.
- peroxisome
- very long-chain fatty acids
- beta-oxidation
1. PPAR
Peroxisome proliferation activating nuclear receptors (PPARs), members of the steroid hormone nuclear receptor ligand-dependent transcription factor superfamily, can regulate peroxisome proliferation by regulating the peroxisome proliferation β-oxidation process [1][39]. PPARs have three subunits, namely PPARα, PPARβ/δ, and PPARγ (Figure 13), which differ in tissue distribution, ligand affinity, and target genes. Surprisingly, they are all related to peroxisomal β-oxidative metabolism [2][40]. Activation of PPARα promotes fatty acid entry into peroxisomes, and PPARα participates in the regulation of energy homeostasis. By contrast, activation of PPARγ causes insulin sensitization and enhances glucose metabolism, whereas activation of PPARβ/δ enhances fatty acid oxidation (FAO).
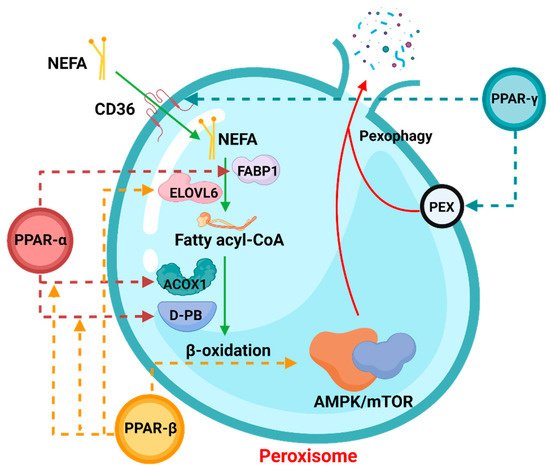
Figure 13.
The role of PPARs in peroxisomal β-oxidation.
In the process of peroxisomal β-oxidation, PPARs are actively involved in various processes. The dotted arrows in the figure indicate that the corresponding PPAR members participate in the regulation of the process. Among them, NEFA: non-esterified fatty acid, FABP1: fatty acid binding protein 1, ELOVL6: ELOVL Fatty Acid Elongase 6, ACOX1: Acyl-CoA Oxidase 1. The process involving pexophagy was complicated, and “PEX” was only a general reference. For details, please refer to Section 3.3.
1.1. PPARα
The effect of peroxisome proliferators on the synergistic induction of the peroxisomal β-oxidation system enzymes has been well established, which is regulated by PPARα. As a lipid sensor, PPARα can coordinate and promote the expression of a large number of target genes in the process of FAO, especially in starvation or high-fat diet conditions [3][41]. Studies have shown that PPARα induces the translation of downstream genes under the stimulation of long-chain fatty acids (VLCFAs): it promotes the proliferation of peroxisomes, and it specifically upregulates the expression of peroxisomal β-oxidative proteins and coenzymes [4][42]. At present, research on PPARα mainly focuses on its targeted ligands. For example, synthetic lipid-lowering drugs have been shown to activate PPARα and then regulate the peroxisomal β-oxidation [5][43]. And in the ruminants, some studies have found that PPARα has different expression levels in the mammary between the dry and lactation period [6][7][19,44]. Further, the study also showed that PPARα plays an important role in regulating milk fat synthesis in ruminants [8][45]. In addition to this, PPARα is also involved in the regulation of fat in the liver [9][10][46,47]. Interestingly, PPARα also plays an important role in ruminant reproduction [11][12][48,49], but it is not known whether this role is related to peroxisomal β-oxidation, which is a direction worth exploring. Simply put, regardless of whether the drugs are endogenous or exogenous ligands, their activation mechanism to bind the PPARα further increases transcriptional activity.
1.2. PPARγ
As one of the key transcriptional regulators of adipocyte differentiation, PPARγ also plays an important role in mediating peroxisomal β-oxidation and lipid metabolisms [13][50]. Studies have shown that a variety of genes involved in fatty acid (FA) transport and metabolism are regulated by PPARγ at the transcriptional level, such as FA translocases, implying that PPARγ can stimulate peroxidase by increasing the expression of FA transporters and FA transportases [14][15][51,52]. The initiation of β-oxidation in vivo has also been proven. By knocking out PPARγ in mice, it was found that they had obvious lipid metabolism disorders [16][53].
In dairy cows, PPARγ plays an important role which is the critical mediator of lipogenesis [17][54]. One of the significant roles it plays in dairy cows from the point of view of economic interest is controlling the synthesis of milk fat in dairy cows, not only in mitochondria but also with ELVOL participating in the regulation of peroxisomal β-oxidation. PPARγ expression increases during changes in the lactation period [18][19][55,56], thereby regulating peroxisomal β-oxidation to prevent metabolic stress. Research further confirmed the importance of PPARγ in regulating milk fat production [20][21][57,58]. Similarly, as milk-producing ruminants, goats and sheep are also regulated by PPARγ. Many studies indicate that PPARγ is involved in adipocyte differentiation and adipogenesis in sheep/goats [22][23][24][25][26][59,60,61,62,63], though, in addition to fat regulation, PPARγ was previously thought to be involved in the regulation of hormones in goats and sheep [27][28][64,65]. However, there has been some relevant research in recent years. Some studies have pointed out that the role of PPARγ and hormones is only a servo-assist mechanism [29][66]. In Up to nowthe author’s opinion, there is no obvious evidence to prove either statement; thus, this is also a worthy question.
1.3. PPARβ/δ
Compared with PPARα and PPARγ, PPARβ/δ is more widely distributed in various tissues in vivo. Aline et al. showed that PPARβ/δ could participate in the activation of FAO in BATs, but genes involved in processes such as lipogenesis were not significantly correlated [30][67]. This demonstrates that PPARβ/δ is a thermogenic transcription factor in vivo, which bears great resemblance to the purpose of peroxisomal β-oxidation. Furthermore, Tong et al. studied PPARβ/δ-induced autophagy, although the study assessed expression changes in mitochondria and revealed the PPARβ/δ-AMPK/mTOR pathway by searching for a signaling pathway (mTOR, one of the important factors of peroxisomal β-oxidation), which further demonstrated that PPARβ/δ has a regulatory effect on peroxisomal β-oxidation [31][68]. Recent studies have shown that PPARβ/δ is indispensable for the upregulation of autophagic behavior [5][32][33][34][35][43,69,70,71,72], making the relationship between PPARβ/δ and peroxisomal β-oxidation more explicit. When the body is under certain conditions, PPARβ/δ frees more VLCFAs by upregulating cellular autophagy, which in turn regulates the initiation and enhancement of peroxisomal β-oxidation. However, a lot of research is still needed to confirm what certain conditions are and whether this speculation is correct.
Regardless of which subunit of PPARs is involved in the regulation of peroxisomal β-oxidation, as PPAR is often reported to be associated with the occurrence of diseases like type 2 diabetes, PPAR-associated peroxisomal β-oxidation changes may also be involved. A study pointed out that inhibition of peroxisome biosynthesis can interrupt β-oxidation through the action of PPAR, thereby effectively preventing the occurrence of type 2 diabetes [36][13]. Nevertheless, the molecular mechanism underlying this phenomenon has not been elucidated. In addition, the prevalence of some diseases is different by gender [37][73], and many experiments also show that PPARs are different in function by gender. PPARα expression is more abundant in native and activated male T cells than in female cells [38][39][74,75], suggesting that PPARα has a more substantial role in male T cells than in female T cells. Likewise, multiple studies have shown that the male hormone androgen has been suggested to influence the expression of PPARα in male T cells [40][41][42][76,77,78]. A study of sex-specific differences in the role of PPARγ in T cell survival has shown that male PPARγ-deficient T cells have increased apoptosis and contain a greater proportion of apoptotic cells than female PPARγ-deficient T cells [43][79]. Some studies have also validated similar conclusions, suggesting that PPARγ plays an important role in T cell survival [44][45][46][47][80,81,82,83]. Although more convincing data are needed to resolve this discrepancy, PPARγ may act as a survival factor in female T cells. Unlike others, the regulatory role of PPARβ/δ in T cells has not been well studied, but sex-specific differences in PPARβ/δ regulation should be considered in future studies.
2. PGC-1α
Peroxisome proliferator-activated receptor-gamma coactivator-1α (PGC-1α) is a powerful transcriptional coactivator that regulates a broad range of physiological and energy homeostasis responses at the transcriptional level in diverse mammalian tissues. In the past, scholars focused more on the regulatory mechanism of PGC-1α in controlling mitochondrial function [48][49][84,85]. In 2010, Bagattin et al. found that PGC-1α coordinates not only mitochondrial remodeling but also peroxisome specialization and biogenesis [50][86], which stimulated an upsurge in the function study of peroxisome PGC-1α. Moreover, Huang et al. studied peroxisomes in human skeletal muscle cells. They found that overexpression of PGC-1α induced the expression of AOCX1, and that the levels of some proteins associated with peroxisome activity also substantially increased [51][87]. In addition, some studies have also demonstrated the importance of PGC-1α in peroxisomes [52][53][88,89].
In conclusion, the regulatory relationship between PGC-1α and peroxisomal β-oxidation deserves further research because of the critical functionality of PGC-1α, and itwe is wondered whether PGC-1α could be used as a novel chemical modulator for the treatment of Zellweger syndrome symptoms and other diseases. PGC-1α has not been studied much in ruminants and has mostly focused on regulating fatty acids [54][55][90,91]. But there is one direction worth mentioning. Zhou et al. found that PGC-1α seems to play a role in the skeletal muscles of goats, and it can also maintain metabolic rhythm through the phosphorylation of upstream regulators [56][92]. ItWe was sspeculated that this is related to peroxisomal β-oxidation.
3. PEX
In 1996, the term peroxin was coined for proteins in peroxisome biogenesis, including peroxisomal matrix protein import, membrane biogenesis, peroxisome proliferation, and peroxisome inheritance [57][93]. Peroxins are encoded by PEX genes, also known as PEX proteins. To date, 37 PEX proteins have been discovered and studied. Some are highly conserved, while others only occur in a limited number of species, such as PEX17, which only distributes in Fungi, and PEX35, which only distributes in Saccharomycetaceae [58][94]. Most PEX proteins have been shown to play a significant role in peroxisomal β-oxidation (Figure 24).
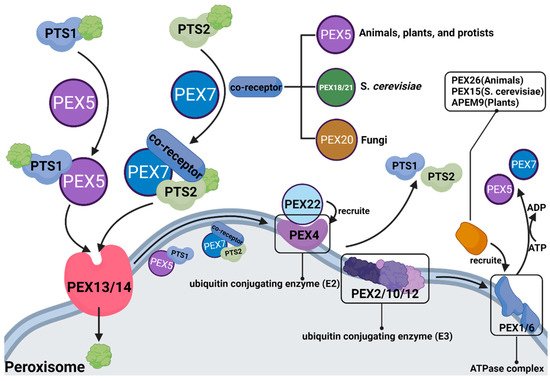
Figure 24.
The role of different PEX proteins in peroxisomal β-oxidation.
PEX5 is the most commonly studied object in eukaryotes. As mentioned above, peroxisomes are very sensitive to reactive oxygen species (ROS)ROS, whereas multiple organelles in the internal environment can produce ROS. Consequently, peroxisomes themselves are susceptible to the influences of other metabolic processes and become dysfunctional. A study found that PEX5 can respond to the expression of ROS and induce cell autophagy after ubiquitination to prevent damage from excessive or defective peroxisomes from cellular β-oxidation [59][95].
More research on PEX has focused on diseases caused by mutation or dysfunction of the PEX gene, including peroxisome biogenesis disorders in the Zellweger spectrum (PBD-ZSD) and rhizomelic chondrodysplasia punctata (RCDP). Studies have pointed out that PBD-ZSD is caused by PEX gene mutation that results in insufficient peroxisomal β-oxidation, which increases levels of VLCFAs in plasma and cells. However, so far, there is no treatment for PBD-ZSD, and researchers have tried to treat it from the perspective of autophagy, but the effect is not ideal [60][96]. Does this imply that there are unknown mechanisms other than autophagy regulating the relationship between PEX and β-oxidation? It reminds us that long-term neglect makes it impossible for us to have precise treatment for diseases caused by peroxisomal β-oxidation disorders, that itwe can’t be found thatfind the therapeutic target, and thus further exploration is urgently needed.
Substances entering peroxisomes for β-oxidation need to form receptor cargo complexes with PTS (PTS: peroxisome targeting sequence, whose role is to locate peroxisomes to ensure accurate transport of carried substances) and PEX, in which PTS1 is transported by PEX5, and PTS2 is transported by PEX7 and coreceptors (coreceptors are PEX5, PEX18/21 and PEX20, depending on the species). After the substance enters the peroxisome, PTS and PEX must be ubiquitinated and recovered.
4. ATP-Binding Cassette (ABC)
In peroxisomes, VLCFAs are mainly introduced as CoA through ABCD1–3 (Figure 35). Studies have shown that peroxisomal β-oxidation is dysfunctional after the deletion of ABCD1/ALDP in vivo, resulting in the expression levels of VLCFAs in both plasma and tissues being increased [61][62][97,98]. ABCD1 and ABCD2 share a high degree of sequence homology, except that ABCD2 plays a central role in the metabolism of monounsaturated and polyunsaturated VLCFAs, rather than saturated VLCFAs, and may be involved in the regulation of oxidative stress and DHA synthesis [63][99]. ABCD3/PMP70 plays a major role in transporting 2-methylacyl-CoA esters [64][100]. In addition, despite the distinct functions of peroxisomal ABC transporters, in vitro and in vivo studies have clearly identified that there is at least partial functional redundancy between these transporters [65][66][101,102]. In ruminants, ABC is equally important. Ahmad et al. found that the expression of ABC varies significantly among different milk yields by RNA-seq [67][103]. Since the expression of ABC is also different between different breeds of cattle, the difference in immunity may also be related to ABC. And Lopez et al. further verified this conclusion, ABC is indeed related to the immune system of cattle [68][104]. Although this conclusion was obtained by RNA-seq, it wase speculated that this is caused by the different content of VLCFAs in vivo and the autophagy function of peroxisome; the latest research also suggests that ABC is involved in intracellular cholesterol-mediated autophagy [69][105]. In addition, the role of ABC was also found in the reproductive system of sheep [70][106], revealing that ABC may be related to the degeneration of germ cells. Although researchers have paid more attention to the transport function of ABCD, itwe would like to be eemphasized that the X-linked adrenoleukodystrophy (X-ALD) remains a matter of concern because it lacks peroxisomal β-oxidation caused by ABCD deletion. At present, the only effective treatment is hematopoietic stem cell transplantation (HSCT), but the risk of death remains high. Some studies have pointed out that the β-oxidation defects can be restored by overexpressing ABCD1/2 in cells, whereas the understanding of its mechanism is still incomplete [71][72][73][28,30,35]. As mentioned above, a comprehensive understanding of peroxisomal β-oxidation is urgently needed.
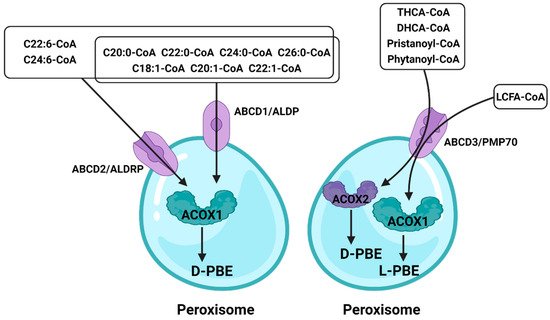
Figure 35.
The role of the ATP-binding cassette in peroxisomal β-oxidation.
Different fatty acids enter peroxisome through different ATP-binding cassettes, and ABCD2 has a broader role. Fatty acids that enter peroxisomes through ABCD3 are processed by different enzymes to achieve oxidation results.
5. Others
New regulators involved in the regulation of peroxisomal β-oxidation have constantly been identified. AMPK, for example, promotes FAO by activating the expression of PPARα and ACOX1 [74][75][107,108], whereas mTOR promotes the accumulation of FAs by shutting down β-oxidation [76][77][109,110]. Hence, these are considered the AMPK-mTOR pathway for regulating β-oxidation [78][111]. The NAD-dependent protein lysine deacylases of the sirtuin family regulate various physiological functions, from energy metabolism to stress responses [79][112]. Numerous studies have found that Sirtuin has a variety of catalytic activities [80][113]. These pleiotropic enzymatic activities give sirtuins their far-reaching functions in maintaining genome integrity, regulating metabolism homeostasis, and promoting organismal longevity. And a recent study found that sirtuin 5 (SIRT5) functions similarly to mTOR, which shuts down peroxisomal β-oxidation by inhibiting the activity of ACOX1 [81][114]. Other reports have studied CoA in the process of β-oxidation and found that Nudt7 and Nudt19 in the Nudt superfamily can exert positive effects on CoA substrates in certain metabolic processes to promote the metabolic process [82][83][115,116]. With the upsurge of research on noncoding RNAs (ncRNAs), some play positive or inhibitory roles in peroxisomal β-oxidation, such as miR-222, miR-25-3p, circ_0005379, and others [84][85][86][87][88][89][117,118,119,120,121,122], which mainly target the rate-limiting enzyme ACOX1. Recently, Li et al. analyzed the ACOX1 transcript and found that miR-532-3P could regulate the expression of ACOX1 by targeting the complementary sequence in the 3’-UTR, thereby participating in lipid metabolism [84][117].
Moreover, other enzymes involved in peroxisomal β-oxidation were also regulated. Several studies have demonstrated that phosphatidylserine (PS) can bind to D-bifunctional protein (D-BP) and localize to peroxisomes, implying that PS can also affect the β-oxidation process [90][123]. It is worth noting that some studies have pointed out that acetylation is important for the normal functioning of D-BP [91][92][124,125], but related studies are not common; thus, itwe is unknowndo not know which factors affect its acetylation process. Another key regulatory enzyme of peroxisomal β-oxidation, Acetyl-CoA acyltransferase 1 (ACAA1), has been extensively studied. In the breeding of livestock and poultry, researchers often explore its effects on adipocytes; for example, in goats and sheep, ACAA1 is involved in regulating adipogenesis. Studies have shown that ACAA1 deficiency increased lipid accumulation and the triglyceride content and promoted sheep preadipocyte differentiation [93][126]. At the same time, it washe study also proved that a a regulatory relationship between ACAA1 and PPARγ. Although it dthe study does not point out the relationship between such a phenomenon and peroxisomal β-oxidation, it is not difficult to speculate that peroxisomal β-oxidation plays an important role. And in humans or model animals, researchers have focused on the diseases caused by it [94][127]. At present, it is generally believed that the expression of PPAR affects the expression of ACCA1, which in turn affects the process of β-oxidation.