Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by François Michel Carlier and Version 2 by Lindsay Dong.
Chronic obstructive pulmonary disease (COPD), asthma and cystic fibrosis (CF) are distinct respiratory diseases that share features such as the obstruction of small airways and disease flare-ups that are called exacerbations and are often caused by infections. Along the airway epithelium, immunoglobulin (Ig) A contributes to first line mucosal protection against inhaled particles and pathogens. Dimeric IgA produced by mucosal plasma cells is transported towards the apical pole of airway epithelial cells by the polymeric Ig receptor (pIgR), where it is released as secretory IgA. Secretory IgA mediates immune exclusion and promotes the clearance of pathogens from the airway surface by inhibiting their adherence to the epithelium.
- immunoglobulin A
- mucosal immunity
- COPD
1. Introduction
Each breath carries thousands of particles towards the airways, constituting many potential threats to lung integrity. Airway mucosal immunity provides both innate and adaptive responses against these inhaled agents, providing inflammatory responses against harmful antigens and tolerogenic mechanisms towards innocuous ones. Failure to these duties may lead to increased antigen penetration and recurrent infections or exaggerated immune responses towards harmless antigens, both situations potentially resulting in chronic airway inflammation. Immunoglobulin (Ig) A represents the predominant Ig at mucosal surfaces in humans [1], where it predominantly lays in its secretory form (S-IgA). Carried towards the airway lumen by its epithelial transporter, the polymeric Ig receptor (pIgR), S-IgA plays a crucial role in the immune exclusion of inhaled pathogens while IgA also regulates immune cells residing in or attracted to mucosal tissues [2].
2. The Mucosal S-IgA System in Homeostasis
The mucosal barrier system is constitutively made of several components. The epithelial layer and the intercellular junctions, with the mucus layer on top, and the lamina propria underneath [3][4][13,14], form a physical barrier against exogenous antigens and pathogens. Mucosal antimicrobial molecules, cellular innate immunity and mucosal adaptative immunity are complementary lines of defence against aggressions [5][15]. In the bronchi, the epithelium is pseudostratified, and composed by numerous cell types whose proportions are tightly controlled [6][7][16,17]. Most of the bronchial airway epithelium consists of ciliated and goblet cells that together constitute the mucociliary elevator clearing particulates and other irritants out of the airspaces. As well as these cell types, the airway epithelium also comprises basal, club and neuroendocrine cells, along with rare ionocytes [8][18]. Ciliated cells are prominent and represent more than 50% of all airway epithelial cells. They possess around 300 cilia whose synchronized beating pushes the mucus layer towards the trachea and the larynx [9][19]. Goblet cells, accounting for 5 to 15% of airway epithelial cells in health, produce mucus and are virtually not present in small airways [9][19]. Basal cells are multipotent stem cells that both anchor the epithelium to the underlying lamina propria and drive epithelial homeostasis and orderly regeneration after injury [10][11][20,21]. They represent 5 to 30% of epithelial cells, their proportion decreasing from the trachea down to the respiratory bronchioles [12][22]. Club cells are dome-shaped cells involved in host defence, and represent 20% of epithelial cells in small airways, where they also behave as progenitor cells [13][14][23,24]. Neuroendocrine cells are rare, innervated cells (< 1% of airway epithelial cells) [15][25] that are thought to be involved in oxygen sensing, smooth muscle tonus and immune responses [16][17][26,27]. Finally, recently discovered ionocytes seem to control the airway surface liquid and mucus viscosity [18][28]. Among these cell types, ciliated, goblet and club cells have been robustly shown to express the polymeric immunoglobulin receptor (pIgR) and therefore participate in the epithelial transcytosis of dimeric IgA (d-IgA) towards the apical mucus layer (see below) [19][20][29,30], while recent single cell transcriptomic data and the emergence of the Human Lung Atlas suggest that ionocytes (but not basal and neuroendocrine cells) also express the pIgR [21][31].2.1. Production and Structure of S-IgA and pIgR
IgA constitutes the most prevalent Ig isotype at mucosal sites and the second most prevalent in serum after IgG. Therefore, IgA is the most abundantly produced Ig in the human body, with an average production rate of 66 mg/kg/day [22][23][32,33]. IgA largely mediates the adaptive humoral immune defence at mucosal surfaces [24][25][34,35], while its role in serum remains relatively unexplored. IgA is produced by B cells both in the systemic and mucosal immune systems, the latter being referred to as mucosa-associated lymphoid tissue (MALT) which includes mucus layers, epithelial cells, lymphoid tissues and immune molecules of the mucosal lamina propria [26][36]. Structurally, IgA is found as monomers (m-IgA) or polymers, mainly consisting of dimers (d-IgA), although some larger forms also exist. M-IgA is composed of two light chains κ and λ, common to all types of Ig’s, covalently linked to two specific heavy chains α [27][37]. In serum, IgA represents 6 to 15% of total immunoglobulins [28][38], and predominates as monomers (85% to 90%). Serum IgA mainly originates from the bone marrow, although the spleen and peripheral lymph nodes could contribute to a lower extent [28][38]. In humans, two IgA subclasses exist, namely IgA1 and IgA2. Each of these IgA subtypes can be found in a monomeric, dimeric, or secretory form [29][40]. In bronchial secretions, IgA2 accounts for 30% of total IgA, compared to 10% in plasma [29][40]. B cells that secrete IgA in the airways are antibody-secreting plasma cells that are generated following cognate interactions between T cells and dendritic cells that have taken up and processed a specific antigen [30][31][42,43].2.2. Transcytosis of d-IgA and Functions of S-IgA
pIgR-mediated polymeric Ig transcytosis requires four well-orchestrated steps [32][51]. First, d-IgA (or pentameric IgM) binds to pIgR through a non-covalent interaction between Ig’s J chain and the extracellular part of pIgR. Second, pIgR is internalized in the clathrin-mediated endocytosis process and delivered in basolateral early endosomes, then in common endosomes. Endocytosed pIgR is then either recycled back to the basolateral membrane, or delivered to the apical surface [33][50]. Third, the extracellular part of the receptor (SC, bound to the transported Ig), undergoes endoproteolytic cleavage by a host serine proteinase. Lastly, S-IgA consisting of d-IgA and SC, crosses the mucus layer by diffusion. Of note is that, unbound pIgR may also undergo epithelial transcytosis and proteolytic cleavage, thereby releasing free SC that may be found in mucosal secretions [32][33][50,51]. At mucosal surfaces, S-IgA functions include neutralization, as well as other biological effects through interactions with, and regulation of, immune cells (via specific Fcα receptors) and microbiota components (Figure 1). The first-line mucosal defence is primarily exerted by S-IgA through its binding of soluble or particulate antigens, preventing their adherence to the epithelium. This non-specific immunity process called ‘immune exclusion’ consists of several sub-steps, including agglutination and entrapment as well as clearance at mucosal surfaces [2][34][2,53].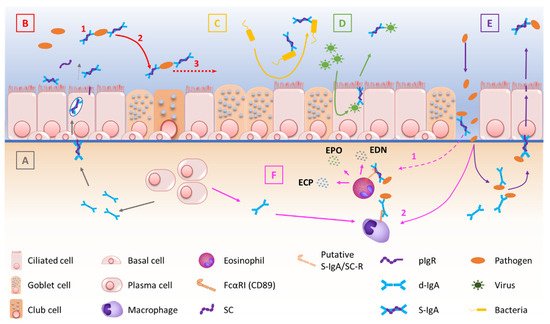
Figure 1. Schematic description of the multiple functions of IgA and the pIgR/S-IgA system at mucosal surfaces. (A) pIgR-mediated endosomal transcytosis of d-IgA, produced by submucosal B cells. (B) S-IgA-driven immune exclusion, including agglutination (1), entrapment of immune complexes (2) and clearance of trapped pathogens (3). (C) Inhibition of bacterial adherence to the mucosal epithelium. (D) Intraepithelial neutralization of penetrating viral antigens. (E) pIgR-mediated elimination of subepithelial immune complexes, after immune exclusion of pathogens by subepithelial d-IgA. (F) (1) S-IgA-induced degranulation of eosinophils resulting from binding of S-IgA to its eosinophil receptor (potentially FcαRI or another receptor such as C-type lectin), occurring possibly after epithelial barrier disruption and releasing eosinophil granule proteins. (2) IgA-induced engagement of phagocytes, enhancing clearance mechanisms.
2.3. Regulation of S-IgA Production
First, the PIGR gene promoter displays binding sites for inflammation-related factors such as IFN regulatory factor 1 (IRF-1), STAT6 and Nuclear Factor (NF)-κB [35][52]. Therefore, host cytokines that activate pathways involving STAT, IRF or NF-κB, such as TNF-α, IFN-ɣ, IL-4 and IL-1, are able to upregulate pIgR expression and d-IgA transepithelial routing [32][35][36][51,52,68]. Depending on the studies, however, exposure to inflammatory stimuli provides divergent results. For instance, IL-4 may stimulate pIgR expression in Calu-3 cell line cultures [37][69], while it inhibits pIgR expression in primary airway epithelial cells [38][70], contributing to pIgR downregulation found in the airway epithelium of asthma patients. A similar dual effect in cell line versus primary cells was observed with TGF-β1, as exogenous exposure of Calu-3 cells to TGF-β1 increases SC production, whereas pIgR production is inhibited by TGF-β1 in primary human bronchial epithelial cells [36][39][68,71]. The molecular substratum for such discrepancies remains unknown. In addition, inflammatory cytokines contribute to pIgR downregulation both in asthma and COPD, while IL-17 conversely upregulates pIgR in Pseudomonas aeruginosa (Pa) infected CF cells [36][38][40][41][68,70,72,73].
Finally, environmental factors may also influence pIgR expression. For instance, pIgR mRNA levels are increased in ex-smokers’ lungs. This increase is however not observed at the protein level, or in situ nor in air/liquid interface cultures of primary human bronchial epithelial cells exposed to cigarette smoke (CS) or derived from smokers [36][42][68,74]. These data suggest that the CS exposure acts as a player in the regulation of pIgR gene expression in vivo, which is further submitted to post-transcriptional modifications that have not been much explored so far. In addition, the microbiota also regulates pIgR through the release of microbe-associated molecular patterns (MAMPs), which control PIGR gene transcription through the activation of Toll-like receptors (TLR) [32][35][51,52].
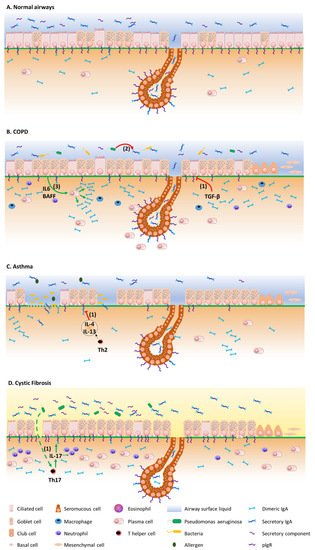
3. The Mucosal S-IgA System in Airway Disease
Figure 2 summarizes the mechanisms by which the IgA/pIgR system is altered in chronic respiratory diseases. Although resulting from complex physiopathological mechanisms in these diseases, respiratory tract colonization by pathogens and trespassing of the mucosal barrier have been shown to trigger these diseases, demonstrating their contribution to the development of such diseases [43][44][75,76].Figure 2. Overview of the pIgR/IgA system dysregulation mechanisms in chronic respiratory diseases. (A) pIgR/IgA system in airway homeostasis, at the epithelial surface and in submucosal glands. (B) In COPD, TGF-β induces pIgR downregulation at the epithelial surface (1), while pIgR expression is preserved in the submucosal glands. The S-IgA local deficiency relates to the subsequently decreased IgA transcytosis, as well as to S-IgA proteolysis by pathogen-derived proteinases (2), favouring bacterial invasion and innate immune cell infiltration. Subepithelial IgA may accumulate as a result of decreased transepithelial transport and IL-6- and BAFF-driven IgA synthesis (3). Enhanced survival of IgA+ plasma cells around the submucosal glands could contribute to a preserved S-IgA production at this level. (C) In asthma, IL-4/IL-13 may induce pIgR downregulation, also leading to luminal S-IgA deficiency (1). (D) In CF, pIgR is conversely upregulated, along with increased production of IgA and S-IgA in airway tissues and lumen, possibly through chronic infection by Pseudomonas aeruginosa that drives pIgR upregulation through IL-17 (1).
3.1. COPD
The functionality of the IgA/pIgR system in respiratory diseases was first explored in COPD, where the abundant literature now clearly demonstrates its multifaceted alteration [45][77]. Chronic obstructive pulmonary disease (COPD), currently representing the third leading cause of death worldwide [46][3], is mainly due to CS with potential additional contributions of other toxics (biomass, occupational, air pollution) and genetic predisposition [47][78].
In 2001, aour team first showed that pIgR/SC expression is decreased in the epithelium of large and small airways from COPD patients, as compared with both non-smokers and non-COPD smokers. In addition, SC expression inversely correlates with COPD severity and related functional parameters such as FEV1, FVC and MEF50 [48][81]. These observations were later corroborated in a larger cohort demonstrating that pIgR epithelial expression is mainly decreased in the airway epithelium from severe COPD patients, as compared with non-smoker controls, non-COPD smokers and less severe COPD patients, and associated with a persistence of the defect in primary cultures of bronchial epithelial cells from such patients where the mechanism could be shown as involving TGF-β signalling [36][68]. Recently, a genome-wide association study showed that PIGR gene expression was reduced in airway intermediate and ciliated cells from smokers without COPD [49][82]. Interestingly, pIgR downregulation in COPD is more obvious in zones of bronchial epithelial remodelling, such as goblet cell hyperplasia, squamous metaplasia, or incompletely differentiated areas. Accordingly, these specific zones also display S-IgA deficiency [50][83], contributing to localized reduced mucosal immunity, with those areas also exhibiting increased bacterial invasion, macrophage and neutrophil infiltration, as well as NF-κB activation [51][84].