1. Eosinophils Regulate the Intestinal Barrier Function
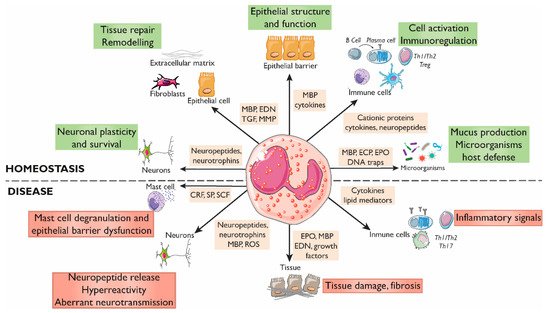
Although the impact of eosinophils on the intestinal epithelium in in vivo human studies is difficult to decipher, data gained from in vitro and pre-clinical models have shown a significant influence of eosinophils on mucosal homeostasis (Figure 12).
Figure 12. Roles of eosinophils in gastrointestinal homeostasis and disease. The interactions between eosinophils and other constituents of the intestinal mucosa (nerves, immune cells, endothelium, stroma, and luminal factors) mediate barrier integrity maintenance or tissue damage through the release of granule mediators. Major basic protein and eosinophil-derived cytokines contribute to barrier function through prostaglandin and ion secretion and mucus production. Eosinophils also impact tissue immuno-regulation and inflammation through cytokines and growth factors that promote adaptive and innate immunity, including a direct effect on mast cells in inflammatory or under stress conditions. Eosinophils also prime B cells for antigen-specific IgM production and sustain long-lived plasma cells. They participate in protection against infections through the release of DNA traps and the stimulation of mucus production through cationic proteins. In addition, eosinophils are also a source of a varied range of proteins and cytokines involved in fibrogenesis and angiogenesis that promote tissue remodeling, repair, and fibrosis. Eosinophils can regulate the function, survival, and development of nerve cells through the release of eosinophil-derived neuropeptides and neurotrophines, MBP, and ROS, and can promote hyperreactivity and aberrant neurotransmission in pathological conditions. CRF, Corticotropin releasing factor; ECP, Eosinophil cationic protein; EDN, Eosinophil-derived neurotoxin; EPO, Eosinophil peroxidase; MBP, Major basic protein; MMP, Matrix metalloprotease; ROS, Reactive oxygen species; SCF, Stem cell factor; SP, Substance P; TGF, Transforming growth factor.
The intestinal epithelium exerts a broad variety of functions ranging from simply being a physical barrier to luminal content to directly promoting active defensive immunity, achieved thanks to the existence of bi-directional communication between intestinal epithelial cells and the whole plethora of mucosal resident immune cells. Eosinophil-induced promotion of intestinal barrier protection seems to depend on the abundance of eosinophils, a low eosinophil–epithelial cell ratio being associated with enhanced barrier integrity
[1][2][3][26,27,28], while a total depletion of intestinal eosinophils is associated with barrier disruption, as experimentally demonstrated by a high-fat diet treatment
[4][29]. A remarkable exception to this pattern is the case of the healthy esophagus, where increased eosinophil numbers in EoE are linked to chronic inflammation and barrier disruption
[5][30]. Notably, the close proximity of eosinophils and intestinal epithelial cells facilitates the crosstalk between these two cell types which may evoke a wide range of effects, ultimately contributing to maintenance of homeostasis or barrier dysfunction.
The interplay between eosinophils and the airway epithelium has been extensively studied, and similar interactions may be expected in the intestinal mucosa. Indeed, a large number of studies have demonstrated that, similar to the airways, intestinal epithelial cells are potent sources of immune-modulating cytokines, chemokines, and growth factors, with large effects on eosinophil recruitment, survival, and activation
[6][7][12,19]. On the other hand, eosinophils can signal to epithelial cells through the generation of reactive oxygen species (ROS) and the release of leukotrienes and other granule-stored mediators, thus augmenting local inflammation and promoting epithelial proliferation
[8][9][31,32]. In EoE, eosinophil-derived mediators have been shown to target esophageal epithelial integrity. More specifically, IL-13 down-regulates the intercellular adhesion molecule desmoglein-1, but not desmoglein-3, leading to altered epithelial barrier function and the promotion of inflammatory responses
[10][11][33,34]. Eosinophils also modulate the secretion of mucus and bactericidal substances contributing to the reinforcement of epithelial barrier protection against infection. In fact, eosinophil-deficient mice show significantly reduced numbers of mucus-secreting goblet cells in the small bowel
[12][35]. Moreover, supernatants derived from activated eosinophils increase mucin levels in human airway epithelial cells
[13][14][36,37], and blocking the eotaxin receptor with anti-CCR3 monoclonal antibody in a mouse model of asthma reduces lung eosinophil recruitment and mucus overproduction
[15][38]. Additionally, eosinophils have the ability to synthesize and rapidly release a web-like complex meshwork of DNA fibers and granule proteins called eosinophil extracellular traps (EETs) in a ROS-dependent manner but independently of eosinophil death, creating a second physical barrier that limits bacterial invasion
[16][39]. However, EETs could also have deleterious effects on intestinal health due to increased production of mucus secretions
[17][40]. Indeed, EETs have been associated with respiratory diseases and a number of other inflammation-associated diseases affecting the skin (atopic dermatitis, urticaria) and the gastrointestinal tract (IBD and EoE)
[17][40].
Additional to the direct effect on the epithelium, eosinophils also contribute to barrier maintenance by promoting tissue remodeling/repair after tissue damage induced by pathogens, toxins, or cell death. A dual role of eosinophils in tissue degradation and repair has been described. In fact, eosinophils release eosinophil-derived neurotoxin (EDN), which promotes fibroblast proliferation
[18][41], and major basic protein (MBP), which synergizes with IL-5 and transforming growth factor beta (TGF-β) to enhance fibroblast expression and secretion of IL-6 and IL-11
[19][42]. In addition, MBP and EDN are potent heparanase inhibitors, which suggests that eosinophils may be involved in proteoglycan degradation prevention and contribute, therefore, to proteoglycan accumulation in fibrotic tissues
[20][43]. Tissue degradation can be sensed by eosinophils which react to promote healing by facilitating epithelial proliferation and acting on the vasculature through the release of vascular endothelial growth factor, fibroblast growth factor, and transforming growth factor-β1 (TGF-β1)
[21][2] in order to recover tissue structure and promote homeostasis.
2. Eosinophils Modulate Intestinal Immune Responses
Eosinophils are a major cellular element of the intestinal mucosa and participate in the regulation of immunity and in the promotion of inflammatory responses (
Figure 2). They are omnipresent at sites of Th2-mediated inflammation, where they can precede or be rapidly recruited to tissue, independently of adaptive immune responses
[22][44]. This suggests that eosinophils themselves can modulate and/or sustain the Th2 character of the local tissue immune microenvironment and also promote Th2 memory cell functions by, for example, releasing cytokines
[23][45]. Beyond Th2 immunity, preformed eosinophil mediators are also related to other immune responses; these include cytokines associated with Th1 (IFN-γ, IL-12), T-regulatory (IL-10, TGF-β), and Th17
[24][25][46,47] activities. In addition, eosinophils have the capacity to initiate and polarize adaptive immune responses. Unlike their blood counterparts, intestinal eosinophils constitutively express on their cell surface antigen presentation markers, including MHC class II, CD80, and activating receptor FcγRIII, suggesting that gut eosinophils may be primed for antigen presentation
[26][27][28][20,48,49]. In addition, eosinophils also promote the initiation of adaptive immune responses through the granule proteins EDN and EPO which are involved in dendritic cell migration, activation, and maturation, leading, therefore, to enhanced antigen-specific Th2 responses
[29][50]. Notably, eosinophils contribute to immunoglobulin production by plasma cells, as shown in animal studies by mediating adjuvant-elicited priming of B cells and optimal antigen-specific early IgM through IL-4
[30][51]. Eosinophils are crucial for class-switch generation and the maintenance of IgA plasma cells in the
lamina propria, mainly through the expression of the A proliferation-inducing ligand (APRIL) and other cytokines, as identified in models of eosinophil-depleted mice
[31][6].
Eosinophil-derived cytokines and chemokines also have effects on innate immune cells, especially on mast cells. Eosinophil–mast cell interactions have been the focus of many hypotheses trying to explain mucosal immune responses (reviewed in
[32][52]). The large amount of evidence of eosinophil and mast cell contribution in the same scenarios, along with the important role mast cells play in allergic and inflammatory diseases also related to the brain–gut axis, suggest an additional eosinophil-derived effector function in immunoregulation within this axis. This potential contribution should no longer be interpreted as a minor interaction based on the release of eosinophil proteins and their inflammatory activity. In fact, eosinophils can promote mast cell growth, survival, and activation by several cytokines and by granule-derived proteins, such as MBP, stem cell factor (SCF), and nerve growth factor (NGF)
[32][33][52,53]. Additionally, mast cells secrete mediators needed for reciprocally activating and promoting the survival of eosinophils
[33][53]. However, mast cells may not be necessarily required for eosinophil survival, as in a mast cell-deficient mouse model of EoE the number of eosinophils remain unaffected
[34][54].
3. Eosinophil–Neuron Interactions
3.1. Innervation of the Gastrointestinal Tract
The gastrointestinal tract is densely populated by two complex networks of neurons (intrinsic innervation) and immune cells that have co-evolved mechanisms to sense and rapidly adapt to the highly dynamic environmental challenges taking place at the intestinal mucosa. The first network is the submucosal plexus or Meissner’s plexus, located in the submucosal region between the circular smooth muscle and the submucosa, which controls glandular secretions, regulates local blood flow, and controls water secretion into the lumen. Fundamental research in different species, such as guinea pigs, rats, and humans, has shown that secretomotor neurons release acetylcholine (ACh) and vasoactive intestinal polypeptide (VIP) which stimulate chloride (Cl-) and water secretion. Nevertheless, the findings in mice are contradictory. In this animal species, the activation of submucosal neurons by electrical field stimulation involves in part the release of Ach and the activation of muscarinic receptors; however, the pharmacological stimulation of these neurons with veratridine does not involve the release of Ach
[35][55]. The second network, called the myenteric plexus or Auerbach plexus, is located between the circular and the longitudinal smooth muscle layers and plays a crucial role in controlling gastrointestinal motility. Excitatory motor neurons synthetize and release ACh as the main neurotransmitter, along with substance P (SP) and other tachykinins. Inhibitory motor neurons synthetize nitric oxide, the main inhibitory neurotransmitter in the upper gastrointestinal tract, and adenosine triphosphate (ATP), the main inhibitory neurotransmitter in the lower gastrointestinal tract. Both plexi of the enteric nervous system (ENS) operate independently but are in turn modulated by the autonomic nervous system via efferent sympathetic and parasympathetic innervation (extrinsic innervation). The cell bodies of these nerves are located in the celiac, superior, and inferior mesenteric ganglions of the sympathetic chain and in the brain, respectively. The central nervous system (CNS) processes sensory information from the different layers of the gastrointestinal tract via the vagal afferent (non-painful physiological stimuli) and spinal afferent nerves (nociceptive stimuli—pain). Gastrointestinal symptoms are triggered basically through the stimulation of chemosensitive nociceptors present in spinal afferent nerves that innervate the
lamina propria and through the activation of mechanosensitive nociceptors present in the longitudinal and circular smooth muscle
[36][56]. The activation of spinal afferent nerves induces the release of sensory neuropeptides, such as SP, CGRP, and NKA.
How the different branches of the extrinsic and intrinsic nervous system crosstalk with innate and adaptive immune cells residing in the gut to jointly coordinate critical physiological functions and responses to challenges has been a matter of intense research. Indeed, functional neuro-immune interactions have been described as playing fundamental roles in intestinal health and disease
[37][57] and there are excellent reviews discussing neuronal crosstalk in several gut immune populations (mainly mast cells, macrophages, and T- and B-cells)
[38][39][9,10].
Like other gut resident immune cells, eosinophils express a wide range of neuropeptides and their receptors that confer on them abilities of interacting with the nervous system, either directly by cell-to-cell contact or indirectly through eosinophil crosstalk with other immune cells residing in the
lamina propria [25][37][47,57]. This eosinophil–neuron communication has been shown to have a bi-directional nature (
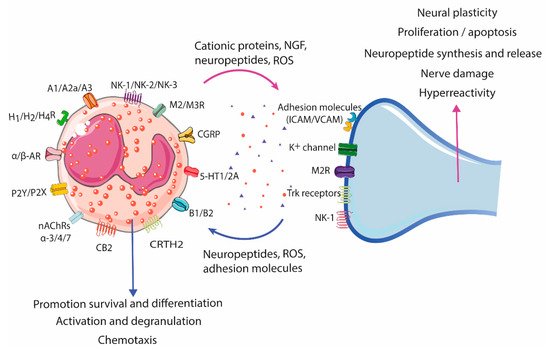
Figure 23); eosinophils can be either the source or the target of the interactions. Indeed, neurons can recruit and activate eosinophils, while eosinophils have been described to show trophic, stimulatory, and inhibitory effects on neurons.
Figure 23. Eosinophil–neuron interaction. Eosinophils and neurons communicate bidirectionally, modulating functions in both cell types. Nerve cells can recruit eosinophils through the release of neuropeptides, cytokines, and chemokines, and activate degranulation through nerve contact facilitated by adhesion molecules (intercellular and neural cell adhesion molecules (ICAMs and NCAMs). The release of bioactive mediators by eosinophils, such as cationic proteins, ROS, and neuropeptides/neurotrophines, results in a variety of neuronal effects, including nerve growth, nerve damage, neuropeptide synthesis and release, and activation and sensitization, leading to hyperreactivity and aberrant neuropeptide release in pathological conditions. A1/A2a/A3, Adenosine receptor; AR, Adrenergic receptor; B1/B2, Bradykinin receptor; CB2, Cannabinoid receptor 2; CGRP, Calcitonin gene-related peptide; CRTH2, Chemoattractant receptor-homologous molecule expressed on Th2 cells; H1/H2/H4R, Histamine receptor H1, H2, H4; ICAM, Intercellular cell adhesion molecule; M2/M3R, Muscarinic receptor 2, 3; nAChRs α-3/-α4/α-7, Nicotinic acetylcholine receptor α-3, -α4, and α-7; NK-1/NK-2/NK-3, Tachykinin receptor 1, 2, 3; P2Y/P2X, Purinergic receptor 2Y, X; ROS, Reactive oxygen species; TrK, Tropomyosin receptor kinase; VCAM, Vascular cell adhesion molecule; 5-HT1/2A, 5-hydroxytryptamine 1, 2A.
3.2. Neural-Induced Recruitment and Activation of Eosinophils by Extrinsic Nerves
Different in vitro and in vivo studies have shown that airway nerves actively recruit and bind eosinophils, promoting a wide range of effects on both cell types. Direct recruitment of eosinophils can happen through eotaxins constitutively expressed and released by parasympathetic efferent nerves, as occurs in the airways during antigenic challenge
[40][41][58,59]. In addition, neurotransmitters and sensory neuropeptides released from the peripheral nerve endings of sensory neurons (extrinsic innervation), such as SP, CCK-8, NKA, and CGRP, can also promote eosinophil recruitment
[42][43][44][45][60,61,62,63]. After recruitment, eosinophils adhere to nerves through cell adhesion molecules (CAMs), which leads to eosinophil activation and degranulation
[43][45][61,63].
After recruitment, eosinophils adhere to nerves through cell adhesion molecules (CAMs), which leads to eosinophil activation and degranulation
[43][45][61,63]. Two major CAM families are involved in eosinophil–nerve binding: the immunoglobulin superfamily of CAMs, which include vascular, intercellular, and neural cell adhesion molecules (VCAM, ICAM, and NCAM); and integrin CAMs, mainly integrin α4β1/VLA-4 and leukocyte function-associated molecule 1 (LFA1). CAM expression on nerves can be constitutive or inducible by pro-inflammatory cytokines and other mediators. Indeed, ICAM expression is induced by TNF-α and IFN-γ causing increased eosinophil adhesion and switching binding preference from VCAM to ICAM in primary cultures of airway parasympathetic nerves
[46][92], an effect that is prevented by treatment with dexamethasone and NFKB inhibitors
[47][93]. Eosinophil engagement to neural adhesion molecules leads to eosinophil activation and ROS production
[48][49][94,95]. Indeed, ICAM and VCAM activation concurrently induce neurite retraction via the generation of tyrosine kinase-dependent ROS and by the p38 MAP kinase pathway
[50][96]. At the same time, neural-derived ROS trigger eosinophil degranulation
[36][56], which makes ROS production a shared event in the eosinophil–nerve bi-directional interaction.
Additional neural stimuli, such as ACh released by peripheral efferent nerves, have been linked to eosinophil chemotaxis and degranulation in experimental models
[51][97] and in atopic asthma patients
[52][98]. It is likely that a similar mechanism exists in the gastrointestinal tract, but further investigations are needed to define the eosinophil–nerve interactions and their contributions to organ functions.
3.3. Neural-Induced Recruitment and Activation of Eosinophils by the ENS
Although the anatomy and function of extrinsic nerves are very different, eosinophil–enteric nerve interactions seem to be similar to those described in the parasympathetic efferent innervation of the lungs. Interestingly, eosinophils are in close proximity to terminal varicosities of excitatory motor neurons expressing SP and choline acetyltransferase (ChAT) but not neuronal nitric oxide synthase (nNOS)
[53][87]. In addition, another cell adhesion molecule, NCAM, has been reported to play a role in eosinophil adhesion to myenteric terminal varicosities in the colonic mucosa of rats undergoing a Th2 response caused by parasitic infection
[54][99]. However, the specific mechanisms underlying eosinophil adhesion to excitatory terminal varicosities during intestinal inflammation are still unknown.
In the gastrointestinal mucosa, SP signaling has been shown to trigger the production and release of corticotropin releasing factor (CRF) in vitro and in experimental chronic restraint stress models
[55][56][79,100]. CRF is a major mediator of stress-induced autonomic, hormonal, and behavioral reflexes that inhibit inflammatory responses at regional levels and influence gut motility and secretion
[57][101] and it has been recently involved in a non-classical non-pro-inflammatory eosinophil activation mechanism, wherein neuromediators selectively induce eosinophil synthesis and release of CRF by piecemeal degranulation
[58][102]. VIP is a key signaling molecule in the neuro-immune network that is secreted by neuronal cells and by different types of immune cells and exerts a wide spectrum of functions. The release of VIP by enteric neurons innervating the intestinal mucosa modulates the epithelial barrier
[59][103]. VIP also regulates the production of both anti- and pro-inflammatory mediators in immune cells
[60][91]. Eosinophils do not express the classical VPAC-1 and VPAC2 but respond to VIP through the Chemoattractant Receptor-Homologous Molecule Expressed on Th2 Cells (CRTH2)
[61][90]. The main effects of VIP on eosinophils are the promotion of chemotaxis and the production of prostaglandin D2, both identified in the context of allergic rhinitis. It may be relevant to determine whether eosinophilia is promoted by VIP released by enteric neurons during intestinal inflammatory conditions, and, additionally, whether eosinophils also impact the intestinal barrier by means of VIP, as it can store and release this neuropeptide
[62][104].