Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 2 by Stanley M Tahara and Version 3 by Dean Liu.
Cancer contains tumor-initiating stem-like cells (TICs) that are resistant to therapies. Experimental evidence indicates that hepatocellular carcinoma (HCC) and TIC development are influenced by permissive conditions in response to changes in gut microbiota.
- cancer stem cell (CSC)
- tumor-initiating stem-like cells (TICs)
- hepatocellular carcinoma (HCC)
1. Introduction
Treatment options for HCC are limited. The 3- or 5-year survival rate of HCC is 13–21% and 5%, respectively, without any curative treatment in advanced countries such as the U.S. [1][2][3][4][5][1,2,3,4,5]. The incidence rate of extrahepatic metastasis is 13% at 5 years [6][7][6,7]. Currently liver resection is the only viable option for HCC combined with cirrhosis that is the terminal stage of fibrosis, leading to hyperplasia formation [8]. Currently, only 10–23% of HCC patients are candidates for surgery [9][10][11][9,10,11]. Thus, HCV-associated HCC remains an incurable malignancy and an urgent unmet medical need. As 40% of HCCs are derived from TICs, TIC-mediated HCC development must be considered as clinically important.
TICs are resistant to conventional chemotherapy and immunotherapy and persist as recurrent tumors or circulating tumor cells (CTC) [12]. TICs express a core pluripotency-associated transcription factor (TF) network [13][14][13,14]. Up to forty percent of HCCs have clonality and are considered to originate from progenitor/stem cells [15][16][17][18][15,16,17,18]. TICs express stemness genes that are also expressed in pluripotent stem cells, including CD133 (Prominin in mice), Wnt/β-catenin, Nanog [19], NOTCH, Hedgehog/SMO, BMI, OCT3/4 [20][21][22][23][24][25][26][27][28][29][30][31][20,21,22,23,24,25,26,27,28,29,30,31], CD44 (cell adhesion molecule), and CD34. CD133+/CD49f+ HCC TICs confer resistance to chemotherapy, which hampers efficacy of therapy in HCC [32]. TICs exhibit a loss of this intrinsic asymmetry, leading to subsequent unchecked expansion of the progenitor cell pool [33][34][35][36][37][38][33,34,35,36,37,38]. Cell-fate-determinant molecule NUMB, and p53-MDM2-associated proteins, are targeted by interacting protein TBC1D15 in TICs [39]. These stemness factors are commonly expressed in TICs and pluripotent stem cells. Stemness factors promote therapy resistance and self-renewal ability.
1.1. Challenge in Targeting of Actionable Mutations
There are no current targeted therapy options for the most prevalent mutations (most are not “actionable”). HCC has only 2.5% of actionable mutations that can be clinically targeted by FDA-approved drugs, while biliary cancer has 45% actionable mutations based on Oncokb.org (Level 3A, Level 3B) and HCC tumor genetics in a TCGA cohort [40]. These indicate many HCC mutations do not have conventional therapeutic targets. Therefore, the role of immunotherapy for the treatment of these diseases is an area of intense investigation [40].
1.2. Molecular Tumor Board (MTB) Review and Actionable Mutations in Liver Cancer
The molecular tumor board (MTB) review can guide choices of therapy for actionable mutations, clarify diagnosis, and identify patients who require germline testing. Prospective clinical sequencing of 10,000 patients revealed the mutational landscape of metastatic cancer [41] Clinical actionability of somatic alterations revealed by MSK-IMPACT was the lowest in HCC mutations at 2.5% [41][42][43][44][45][41,42,43,44,45], while clinical actionability of somatic alterations revealed by MSK-IMPACT showed that 45% of biliary cancer mutations are clinically actionable. The molecular tumor board (MTB) for intrahepatic cholangiocarcinoma (iCCA) shows clinically targetable mutations [46][47][48][46,47,48]. iCCA is a heterogeneous disease with several identifiable genetic driver mutations (i.e., FGFR2-fusions IDH mutations, etc.) [40]. For iCCA, fluorescence in situ hybridization (FISH), DNA/RNA-seq, and immunohistochemistry (IHC) analyses can identify cancer-driver mutations, including IDH1/2, CDK4/67, PRKACA/B, and BRCA1/2. FGFR2 and NYRK fusions, BRAF and IDH1 mutations, and microsatellite instability high (MSI-H)/dMMR (defective mismatch DNA repair) predict responses to targeted/immune therapies [41]. Tumor next-generation sequencing (NGS) should be considered in selected HCC patients with atypical histology/diagnostic features or who may be eligible for clinical trials. HCC classification, cells of origin, genetic and epigenetic abnormalities, molecular alterations, biomarker discovery, and treatments of iCCA have been well characterized [49].
2. Gut-Microbiota-Mediated Immune Regulatory Mechanisms by Immunotherapy
2.1. Bacterial Enzyme Inhibitors Can Be Used for Treatment
Gut microbial produced metabolites can be recognized by host pathogen recognition sensors to promote HCC progression. Metabolism of dietary components by the gut microbiota produces short-chain fatty acids, including other metabolites. When combined with microorganism fragments, these can stimulate the meta-organismal endocrine axis to promote HCC onset and growth. For example, trimethylamine (TMA) produced in the gut promotes ALD [44]. Thus, pharmacological interventions at the level of the gut microbiome should reduce HCC risk. Targeting of the gut microbiota has great potential as a therapeutic modality for many diseases. However, relatively little is known regarding the contribution of commensal bacteria to normal host physiological functions [45]. For example, it was reported that 11 bacterial strains in feces obtained from normal human donors induce CD8 T cells to produce IFN-γ in the intestine in the absence of a generalized inflammatory response dependent on CD103+ DC and MHC class Ia [45]. These 11 strains also improved the efficacy of immune checkpoint inhibitors and aided host suppression against Listeria monocytogenes infection [45]. Thus, these 11 identified strains, which represent low-abundance components of the human microbiome, are potential biotherapeutics [45].2.2. TLR2 Signaling in DCs Promotes Treg Differentiation to Attenuate the Inflammation
TLR2 senses components from bacteria, mycoplasma, fungi, and viruses [47] to activate NF-κB to promote a Th17 cell response to enhance the inflammation response and anti-inflammation responses [48][50][48,109]. Lactobacillus acidophilus stimulates the TLR2 pathway of murine myeloid dendritic cells (mDC) to induce interferon-β (IFN-β), while IL-10 secretion in plasmacytoid DC (pDC) is TLR9 dependent (Figure 13). Bifidobacterium infantis 35624 stimulates the TLR2/TLR6 pathway to increase IL-10 secretion from human DCs. Polysaccharide A of Gram negative bacilli can activate TLR2 and promote the secretion of anti-inflammatory cytokine IL-10 [51][110]. These diverse immune responses depend on the appropriate co-receptor and microenvironment [48].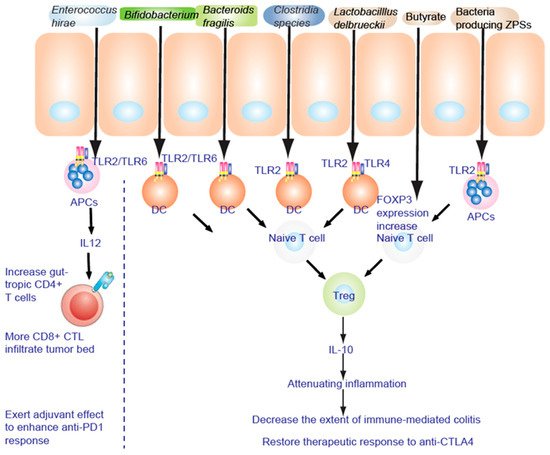
Figure 13. TLR2 is necessary to alleviate the inflammatory response. Fructo-oligosaccharide and inulin are considered as prebiotics, affecting IECs to be hyporesponsive to activation of NF-κB and MAPK induced by pathogens. NF-κB and MAPK reduce the inflammatory response to lipopolysaccharide (LPS).