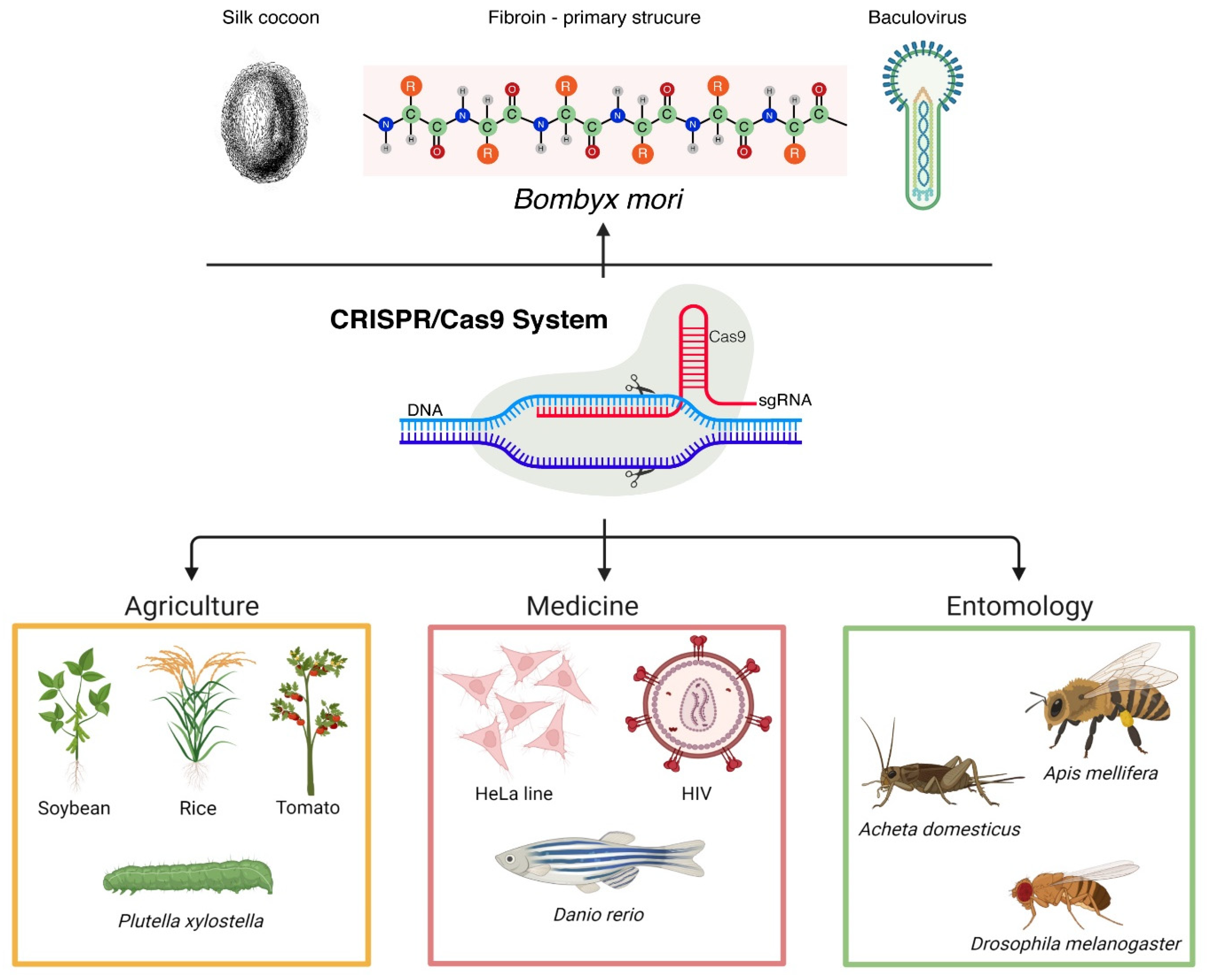
CRISPR (clustered regularly interspaced short palindromic repeats)-Cas (CRISPR-associated) represents a powerful genome editing technology that revolutionized in a short period of time numerous natural sciences branches. Therefore, extraordinary progress was made in various fields, such as entomology or biotechnology.
- CRISPR-Cas
- genome engineering
- insect biotechnology
- applied biotechnology
1. Introduction
2. The CRISPR-Cas System
2.1. The CRISPR-Cas Complex Role in the Immunity System
2.2. The CRISPR-Cas System as a Genome Editing Tool
2. The CRISPR-Cas System
CRISPR-Cas is one of the key methods employed by many molecular biology scientific laboratories. Since its first description [7], genome editing focused research was implemented by countless research groups [8][9][10].2.1. The CRISPR-Cas Complex Role in the Immunity System
When investigating the iap gene product in the opportunistic pathogen Escherichia coli, Ishino et al. (1987) [11] observed an atypical structure, specifically the repetition of several homologous sequences. Later, this type of structure was observed in various bacterial, as well as archaeal strains [12][13]. Subsequently, these repetitive sequences were linked with exogenous genetic material, and following several years, their assembling mechanism and function were elucidated [14]. This type of sequence can be placed on the chromosomal DNA, but it can also be found on the plasmid DNA [15]. The scientists demonstrated that CRISPR-Cas, which is present in one-third of bacteria and nearly in all archaea, has a key role in host’s adaptive immunity. It protects the organism against various intruders, such as viruses, but it also offers protection against other mobile genetic elements, such as transposons or plasmids [16]. The CRISPR-Cas system structure includes three main components, i.e., the CRISPR arrays, the associated Cas proteins, and the leader nucleotide sequence. The first genetic component, the CRISPR locus, is characterized by identical repeats structures (21–37 bp) that are highly conserved and the spacer sequences that are acquired fragments of invader’s nucleic acid material. The CRISPR array is located downstream from cas genes. The latter encodes for Cas proteins that are crucial to the immune reaction [17]. Initially, only four distinct Cas proteins (1–4) were reported, due to the rapid evolution of biological sciences; currently, there numerous Cas proteins have been described [18][19], Cas1 being the most analyzed [20]. CRISPR-Cas has a great adaptability, with host-related specificities; thus, it exhibits a significant diversity. The varying feature is defined by the CRISPR array and the cas gene sequences. The classification of these types of systems is based on the signature Cas proteins. Currently, there are two major classes of CRISPR-Cas systems, each also di-vided in several groups [21]. Regarding the leader nucleotide sequence, it has been shown that it has a key role by carrying the essential promoter sequences for the transcription of CRISPR loci. Besides the promoter, the leader contains specific signals that are crucial for the adaptation stage from the first phase of CRISPR-Cas activation [22]. The adaptation is the first functional stage of the CRISPR-Cas mechanism, during which the foreign nucleic acid is recognized by several Cas proteins [23] and consequently integrated next to a leader sequence. Through this mechanism, in evolution, the spacers are arranged chronologically, and this feature helps bacteria and archaea to enhance their protection against the genetic material of the latest foreign encounter [24]. Each new acquired spacer is accompanied by a repeat sequence; therefore, the CRISPR array expands with every invasion [25]. The CRISPR array is transcribed in the second step, specifically in the biogenesis phase [26]. First of all, it is being transcribed into a precursor CRISPR RNA (crRNA). At the end of this phase, there are numerous mature crRNAs molecules, resulting from the action of RNase III that process the precursor crRNA. Each crRNA includes a spacer and a repeat sequence [24][27][28]. The last step of the system’s mechanism is the interference phase. It involves the degradation of the foreign nucleic acid, by targeting and cleaving it [29]. The products of the biogenesis phase, the crRNAs, act like guides for targeting the invader, which is then cleaved following a Cas proteins cascade that act like molecular scissors [30].2.2. The CRISPR-Cas System as a Genome Editing Tool
When it comes to leading tools in genetic engineering, the CRISPR-Cas system can be considered the foremost instrument. After elucidating its function in various organisms, scientists aimed to exploit its versatility, in order to overcome the disadvantages of other available genome editing tools [31]. Even if scientific studies still report the use of ZFNs and TALENs techniques as editing tools, the CRISPR-Cas system is the most effective genome editing instrument, standing on top in regards to efficiency, cost-effectiveness, and the relative simplicity of use [32] (Traits | TALEN | ZFN | CRISPR-Cas | References | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
Origin | Prokaryotic | Eukaryotic | Prokaryotic |
[41] |
[34] |
|||||||||
Efficiency (%) | 76 | 12 | 81 |
[42] |
[35] |
|||||||||
Specificity | Moderate | Low | High | |||||||||||
Target site recognition | Any site | Any site | Pam motif (NGG) required |
[43] |
[36] |
|||||||||
Multiplex potential | Low | Low | High | |||||||||||
Processing time | Time consuming | Time consuming | Short |
[45] |
[38] |
|||||||||
Methylation sensitive | Sensitive | Sensitive | Not sensitive |
[42] |
[35] |
|||||||||
Engineering feasibility | Moderate/High | Moderate | Moderate/High | |||||||||||
Dimerization required | Yes | Yes | No |
[44] |
[37] |
|||||||||
Cost effectiveness | Moderate | No | Yes |