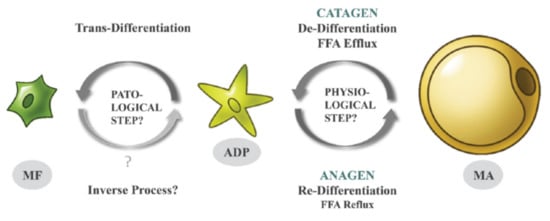
Adipocytes from the superficial layer of subcutaneous adipose tissue undergo cyclic de- and re-differentiation, which can significantly influence the development of skin inflammation under different cutaneous conditions. This inflammation can be connected with local loading of the reticular dermis with lipids released due to de-differentiation of adipocytes during the catagen phase of the hair follicle cycle. Alternatively, the inflammation parallels a widespread release of cathelicidin, which typically takes place in the anagen phase (especially in the presence of pathogens). Additionally, trans-differentiation of dermal adipocytes into myofibroblasts, which can occur under some pathological conditions, can be responsible for the development of collateral scarring in acne.
1. Introduction
Dermal white adipose tissue (dWAT) is a newly recognized fat depot located in the superficial hypodermis [1][2]. Recent findings have revealed that dermal adipocytes are phenotypically different from their anatomically nearby subcutaneous counterparts and demonstrate diverse rapid transformations in response to some external and internal factors. These cells contribute in substantial ways to a variety of processes. This includes cutaneous events, such as hair follicle (HF) cycling, wound healing, and the innate immune response (mainly through the production of antimicrobial peptides); additionally, researchers suggested that dermal adipocytes are involved in scarring and skin aging [1]. One unique property of murine dWAT is the remarkable oscillation of its volume during the hair follicle cycle; it strongly decreases in catagen and completely restores in anagen [2][3][4]. The physiological impact and underlying reasons for these oscillations have been a topic of intensive discussion in the recent past. Dermal white adipose tissue (dWAT) is a newly recognized fat depot located in the superficial hypodermis [1,2]. Recent findings have revealed that dermal adipocytes are phenotypically different from their anatomically nearby subcutaneous counterparts and demonstrate diverse rapid transformations in response to some external and internal factors. These cells contribute in substantial ways to a variety of processes. This includes cutaneous events, such as hair follicle (HF) cycling, wound healing, and the innate immune response (mainly through the production of antimicrobial peptides); additionally, we suggested that dermal adipocytes are involved in scarring and skin aging [1]. One unique property of murine dWAT is the remarkable oscillation of its volume during the hair follicle cycle; it strongly decreases in catagen and completely restores in anagen [2,3,4]. The physiological impact and underlying reasons for these oscillations have been a topic of intensive discussion in the recent past.
Recently, researchers demonstrated, in murine models, that these oscillations are the result of specific cellular conversions; mature dermal adipocytes de-differentiate in the catagen phase into the much smaller fibroblast-like adipocyte-derived preadipocytes (ADPs) and re-differentiate in anagen back into mature adipocytes (
Recently, we demonstrated, in murine models, that these oscillations are the result of specific cellular conversions; mature dermal adipocytes de-differentiate in the catagen phase into the much smaller fibroblast-like adipocyte-derived preadipocytes (ADPs) and re-differentiate in anagen back into mature adipocytes (
Figure 1) [3][4]. Such physiological processes of de- and re-differentiation of adipocytes repeat during every HF cycle and involve a large number of murine dermal adipocytes. The remaining mature adipocytes in the dWAT layer may be recruited from distinct adipose progenitors that were not subject to such previous cyclic conversions. ) [3,4]. Such physiological processes of de- and re-differentiation of adipocytes repeat during every HF cycle and involve a large number of murine dermal adipocytes. The remaining mature adipocytes in the dWAT layer may be recruited from distinct adipose progenitors that were not subject to such previous cyclic conversions.
Figure 1. De-, re-, and trans-differentiation of dermal adipocytes during the HF cycle [5] (with permission from Experimental Dermatology).
De-, re-, and trans-differentiation of dermal adipocytes during the HF cycle [9] (with permission from Experimental Dermatology).
De-differentiation of dermal adipocytes into ADPs in the catagen is accompanied by a significant reduction of their volumes and leads to loading of the dWAT and reticular dermis with fatty acids [6][7]. Re-differentiation of preadipocytes in anagen leads to a regain of the cell volumes and causes intensive expression of the anti-microbial peptide cathelicidin. This expression can be significantly enhanced in the presence of pathogens [8]. Local loading of the skin with extracellular lipids and cathelicidin can induce a local inflammatory reaction in the affected skin areas. De-differentiation of dermal adipocytes into ADPs in the catagen is accompanied by a significant reduction of their volumes and leads to loading of the dWAT and reticular dermis with fatty acids [5,6]. Re-differentiation of preadipocytes in anagen leads to a regain of the cell volumes and causes intensive expression of the anti-microbial peptide cathelicidin. This expression can be significantly enhanced in the presence of pathogens [7]. Local loading of the skin with extracellular lipids and cathelicidin can induce a local inflammatory reaction in the affected skin areas.
Moreover, under appropriate conditions, ADPs can leave this cycle and transdifferentiate into synthetically highly active myofibroblasts. While the specific signals triggering such trans-differentiation still remain to be elucidated, they do involve stimulation by transforming growth factor beta (TGF-β). This type of cellular change does not occur under healthy conditions and can lead to local cutaneous fibrosis and scarring (
Figure 1). Whereas some authors found that myofibroblasts may re-differentiate into adipose cells [9], this issue will need further investigation. ). Whereas some authors found that myofibroblasts may re-differentiate into adipose cells [8], this issue will need further investigation.
Whereas the cell conversions described in [4] directly affect only the local structure and physiology of dWAT, their effects can be transmitted to the dermis and even the epidermis and can thus be involved in the pathophysiology of different cutaneous conditions.
2. DWAT Layer in Human Skin
dWAT has been extensively investigated and well described in murine skin, whereas the existence of this layer in humans is a matter of controversy [1]. The main problem is the obvious inability to conduct lineage-tracing studies in humans [10]. Another problem is the widespread opinion that there is no anatomical demarcation of dWAT in human skin analogous to the Panniculus carnosus (a layer of striated muscle cells) seen in rodents. Some authors have proposed that this structure is underdeveloped in humans [2]. In contrast, others describe fascial planes composed of fibrous connective tissue and muscular fibers producing a separating layer between the dermis and hypodermis, which can be considered a structure homologous to the Panniculus carnosus. Confusingly, these layers have different designations depending on anatomical location, such as Galea aponeurotica in the head, superficial musculo-aponeurotic system (SMAS) in the face, platysma in the neck, Scarpus fascia in the abdomen, and Colles fascia in the perineum [11].
Nevertheless, several authors have proposed the existence of a similar dermal-associated fat depot in humans and described its unique geometry as a flat superficial layer of subcutis with additional adipose cones penetrating the dermis and located around hair follicles (HFs) [1][2][12]. At the same time, this does not mean that dWAT in rodents and humans is anatomically, physiologically, and metabolically identical [4]. To avoid possible confusion, some authors have proposed to designate dWAT in humans as skin-associated adipose tissue (SAAT) [4] or skin-associated fat (SAF) [13].
A possibility is to use chemical shift magnetic resonance tomography to separate the fat-only and water-only images and thus to assess the dWAT depot in rodents and the SAF depot in humans [13][14]. Applying this method, these authors estimated the thickness of the SAF as a body-wide depot to be about 10.05 ± 2.45 mm (females) and 6.27 ± 1.89 mm (males) with a high inter-individual variability of thicknesses. A similar dimorphism was earlier observed in murine dWAT [15] and later confirmed in [13]. No significant correlation was found between the thickness of this layer and the age of the individuals. In humans, the thickness of SAF is independent of the body mass index (BMI) in males and nearly independent of this parameter in females [13], whereas in murine models, dWAT thickness positively correlates with BMI [4]. Interestingly, while adipocytes from dWAT are resistant to β-adrenergic-induced lipolysis [4][13], adipocytes from SAF were found to be highly lipolytic [13].
Combined, there is overwhelming evidence that humans have a skin-associated adipose tissue depot similar to dWAT in rodents. Given that adipocytes from dWAT are involved in various important physiological and pathological processes in the skin, researchers assume that the same is correct for skin-associated adipocytes in humans.
3. Connection of Acne and Psoriasis to the HF Cycle and High Fat Diet
3.1. Acne vulgaris
Acne vulgaris is the most common inflammatory skin disorder with a high prevalence in adolescence, and it is spatially localized to the small skin areas adjacent to pilosebaceous units. The pathophysiology of acne is connected to etiopathogenic factors, such as hyperseborrhea, follicular hyperkeratinization, and the pathogenic behavior of the gram-positive bacterium C. acnes [16]. Hyperseborrhea was traditionally linked to increased sebum excretion, whereas follicular hyperkeratinization was explained by the abnormal differentiation and desquamation of keratinocytes. The pathogenic behavior of C. acnes was mainly explained by the uncontrolled proliferation of these bacteria [5]. Remarkably, perifollicular inflammation typical of acne is found just in the earliest stages of the development of acne lesions [17], which means that Acne vulgaris should be considered a primary inflammatory skin disorder.
The relative contribution of exposure to high-fat diets (HFDs) toward acne is still a matter of controversy. In fact, a recent report argued that adiposity in adolescence is inversely related to acne [18]. At the same time, the mammalian target of rapamycin complex 1 (mTORC1) expression, known to be regulated by HFD, growth factors, and stress and critically involved in the induction of lipolysis in adipocytes, is significantly increased in acne lesions compared to non-lesional skin [19]. HFD also efficiently induces the expansion of dWAT [4]. This suggests that HFD can theoretically influence acne not through a systemic impact but rather through the local modification of the adjacent dWAT. However, other factors cannot be excluded.
Acne vulgaris is not only spatially connected to pilosebaceous units, but it is also strongly dependent on the HF cycle. The initiation of acne lesions was historically assigned to the catagen/telogen phases of the HF cycle [20]. At the same time, there is no known HF cycle-dependent change in C. acnes behavior. Moreover, whereas the sebaceous gland indeed demonstrates varied sebum production in different HF phases, this secretion is normally lower in catagen than in anagen, which argues against predominant production of acne lesions during catagen/telogen phases. This paradox is difficult to explain in the frames of the classical acne pathophysiology, primarily connecting perifollicular inflammation in acne lesions with hyperseborrhea and activity of C. acnes. On the other hand, researchers can explain the phenomenon through temporarily restricted loading of perifollicular areas with free fatty acids released during de-differentiation of mature adipocytes into ADPs in the catagen phase of the HF cycle (
2. DWAT Layer in Human Skin
dWAT has been extensively investigated and well described in murine skin, whereas the existence of this layer in humans is a matter of controversy [1]. The main problem is the obvious inability to conduct lineage-tracing studies in humans [10]. Another problem is the widespread opinion that there is no anatomical demarcation of dWAT in human skin analogous to the Panniculus carnosus (a layer of striated muscle cells) seen in rodents. Some authors have proposed that this structure is underdeveloped in humans [2]. In contrast, others describe fascial planes composed of fibrous connective tissue and muscular fibers producing a separating layer between the dermis and hypodermis, which can be considered a structure homologous to the Panniculus carnosus. Confusingly, these layers have different designations depending on anatomical location, such as Galea aponeurotica in the head, superficial musculo-aponeurotic system (SMAS) in the face, platysma in the neck, Scarpus fascia in the abdomen, and Colles fascia in the perineum [11].
Nevertheless, several authors have proposed the existence of a similar dermal-associated fat depot in humans and described its unique geometry as a flat superficial layer of subcutis with additional adipose cones penetrating the dermis and located around hair follicles (HFs) [1,2,12]. At the same time, this does not mean that dWAT in rodents and humans is anatomically, physiologically, and metabolically identical [4]. To avoid possible confusion, some authors have proposed to designate dWAT in humans as skin-associated adipose tissue (SAAT) [4] or skin-associated fat (SAF) [13].
A possibility is to use chemical shift magnetic resonance tomography to separate the fat-only and water-only images and thus to assess the dWAT depot in rodents and the SAF depot in humans [13,14]. Applying this method, these authors estimated the thickness of the SAF as a body-wide depot to be about 10.05 ± 2.45 mm (females) and 6.27 ± 1.89 mm (males) with a high inter-individual variability of thicknesses. A similar dimorphism was earlier observed in murine dWAT [15] and later confirmed in [13]. No significant correlation was found between the thickness of this layer and the age of the individuals. In humans, the thickness of SAF is independent of the body mass index (BMI) in males and nearly independent of this parameter in females [13], whereas in murine models, dWAT thickness positively correlates with BMI [4]. Interestingly, while adipocytes from dWAT are resistant to β-adrenergic-induced lipolysis [4,13], adipocytes from SAF were found to be highly lipolytic [13].
Combined, there is overwhelming evidence that humans have a skin-associated adipose tissue depot similar to dWAT in rodents. Given that adipocytes from dWAT are involved in various important physiological and pathological processes in the skin, we assume that the same is correct for skin-associated adipocytes in humans.
3. Connection of Acne and Psoriasis to the HF Cycle and High Fat Diet
3.1. Acne vulgaris
Acne vulgaris is the most common inflammatory skin disorder with a high prevalence in adolescence, and it is spatially localized to the small skin areas adjacent to pilosebaceous units. The pathophysiology of acne is connected to etiopathogenic factors, such as hyperseborrhea, follicular hyperkeratinization, and the pathogenic behavior of the gram-positive bacterium C. acnes [16]. Hyperseborrhea was traditionally linked to increased sebum excretion, whereas follicular hyperkeratinization was explained by the abnormal differentiation and desquamation of keratinocytes. The pathogenic behavior of C. acnes was mainly explained by the uncontrolled proliferation of these bacteria [9]. Remarkably, perifollicular inflammation typical of acne is found just in the earliest stages of the development of acne lesions [17], which means that Acne vulgaris should be considered a primary inflammatory skin disorder.
The relative contribution of exposure to high-fat diets (HFDs) toward acne is still a matter of controversy. In fact, a recent report argued that adiposity in adolescence is inversely related to acne [18]. At the same time, the mammalian target of rapamycin complex 1 (mTORC1) expression, known to be regulated by HFD, growth factors, and stress and critically involved in the induction of lipolysis in adipocytes, is significantly increased in acne lesions compared to non-lesional skin [19]. HFD also efficiently induces the expansion of dWAT [4]. This suggests that HFD can theoretically influence acne not through a systemic impact but rather through the local modification of the adjacent dWAT. However, other factors cannot be excluded.
Acne vulgaris is not only spatially connected to pilosebaceous units, but it is also strongly dependent on the HF cycle. The initiation of acne lesions was historically assigned to the catagen/telogen phases of the HF cycle [20]. At the same time, there is no known HF cycle-dependent change in C. acnes behavior. Moreover, whereas the sebaceous gland indeed demonstrates varied sebum production in different HF phases, this secretion is normally lower in catagen than in anagen, which argues against predominant production of acne lesions during catagen/telogen phases. This paradox is difficult to explain in the frames of the classical acne pathophysiology, primarily connecting perifollicular inflammation in acne lesions with hyperseborrhea and activity of C. acnes. On the other hand, we can explain the phenomenon through temporarily restricted loading of perifollicular areas with free fatty acids released during de-differentiation of mature adipocytes into ADPs in the catagen phase of the HF cycle ( Figure 1). Moreover, trans-differentiation of ADPs into myofibroblasts can explain the appearance of hypotrophic scars in long-lasting acne lesions.
3.2. Psoriasis vulgaris
Psoriasis is a common chronic autoimmune hyperproliferative skin disorder with a strong genetic predisposition. Whereas the pathophysiology of psoriasis is not fully elucidated, it is widely accepted that it includes a dysregulation of innate and adaptive skin immunity, epidermal hyperproliferation, and inflammation. The clinically relevant skin modifications in psoriasis were initially found in the epidermis and connected to the pathological behavior of keratinocytes. For this reason, psoriasis has long been considered an inflammatory and hyperproliferative disease of the epidermis.
Psoriasis is considered to be independent of HF cycling. However, remarkable correlations between epidermal proliferation in psoriatic lesions and the anagen phase of the HF cycle were first discussed in [21]. These authors proposed that processes underlying both HF cycling and psoriasis share the same “switch-on” mechanisms. HFs within psoriatic scales demonstrate a remarkable shift in the relative distribution of HFs in the different phases; there are almost twice as many telogen HFs in psoriatic skin [22].
Such a modification in the staging of HF distribution points to a partial synchronization of HFs in psoriatic lesions. This is surprising, since human HFs under normal physiological conditions demonstrate highly asynchronous behavior. However, as they argued previously, HF synchronization can take place in some spatially restricted areas [23], which can also lead to synchronized changes in dermal adipocytes.
Obesity demonstrates multiple comorbidities, and psoriasis is one of them [24]. Epidemiological studies highlight that obesity and weight gain are important risk factors for psoriasis [25][26]. On the other hand, weight loss has a positive impact on the severity of the disease [27]. Specifically, abdominal fat is intricately associated with disease predisposition and progression, and the metabolic health of visceral adipose tissue strongly relates to the manifestations of psoriasis. Skin and adipose tissue interact through adipokines. One of these adipokines—adiponectin—is a well-known regulator of insulin sensitivity, which also modulates proliferation and migration of keratinocytes [28] and modifies psoriasiform skin inflammation through suppression of IL-17 production [29]. Consistent with these observations, adiponectin KO mice can demonstrate a severe form of psoriasiform dermatitis [29]. On the other hand, HFD significantly exacerbates early psoriatic skin inflammation independent of obesity. Furthermore, a reduction of dietary saturated fatty acids improves psoriasiform inflammation independently of weight and fat content [30].
Researchers have proposed that it is not the abdominal adipose tissue; rather, it is the dWAT located at the interface between dermis/subcutis that is involved in the pathophysiology of psoriasis [31][32]. There are several direct indications that dWAT undergoes structural modifications strongly associated with the psoriatic phenotype. PUVA treatment (a combination treatment consisting of application of psoralens (light sensitizers) (P) and then exposing the skin to UVA (long wave ultraviolet radiation) of K5.hTGFb1 transgenic mice (a murine model for psoriasis) caused a significant reduction of the dWAT layer beneath the psoriatic lesions, and such a modification correlated with clinical skin improvements in these animals [33]. Sonoelastographic investigation of the fat tissue located beneath the human psoriatic plaques demonstrates that the mechanical properties of the subcutis, beneath these spatially restricted areas, are very different from those in the areas covered by healthy skin. Successful treatment of psoriasis leads to skin improvements and simultaneous normalization of the adjacent fat tissue [34][35].
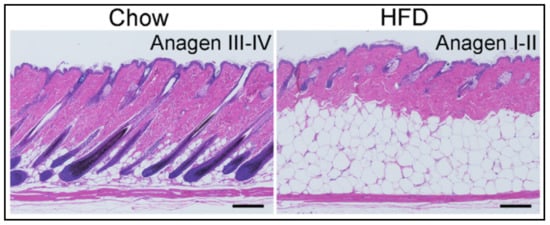
Taking into account the spatially limited distribution of psoriatic lesions and the modifications of dWAT located beneath these lesions [31][32], a critical question is what happens with dWAT during exposure to HFD? As researchers have demonstrated recently, HFD prompts at least two major modifications to dWAT: (1) a dramatic expansion of its volume involving both the processes of hyperplasia and hypertrophy of dermal adipocytes, and (2) a partial synchronization of HFs in the catagen/telogen phases with an impairment of their re-entry into anagen compared to chow-fed controls [4] ( ). Moreover, trans-differentiation of ADPs into myofibroblasts can explain the appearance of hypotrophic scars in long-lasting acne lesions.
3.2. Psoriasis vulgaris
Psoriasis is a common chronic autoimmune hyperproliferative skin disorder with a strong genetic predisposition. Whereas the pathophysiology of psoriasis is not fully elucidated, it is widely accepted that it includes a dysregulation of innate and adaptive skin immunity, epidermal hyperproliferation, and inflammation. The clinically relevant skin modifications in psoriasis were initially found in the epidermis and connected to the pathological behavior of keratinocytes. For this reason, psoriasis has long been considered an inflammatory and hyperproliferative disease of the epidermis.
Psoriasis is considered to be independent of HF cycling. However, remarkable correlations between epidermal proliferation in psoriatic lesions and the anagen phase of the HF cycle were first discussed in [21]. These authors proposed that processes underlying both HF cycling and psoriasis share the same “switch-on” mechanisms. HFs within psoriatic scales demonstrate a remarkable shift in the relative distribution of HFs in the different phases; there are almost twice as many telogen HFs in psoriatic skin [22].
Such a modification in the staging of HF distribution points to a partial synchronization of HFs in psoriatic lesions. This is surprising, since human HFs under normal physiological conditions demonstrate highly asynchronous behavior. However, as we argued previously, HF synchronization can take place in some spatially restricted areas [23], which can also lead to synchronized changes in dermal adipocytes.
Obesity demonstrates multiple comorbidities, and psoriasis is one of them [24]. Epidemiological studies highlight that obesity and weight gain are important risk factors for psoriasis [25,26]. On the other hand, weight loss has a positive impact on the severity of the disease [27]. Specifically, abdominal fat is intricately associated with disease predisposition and progression, and the metabolic health of visceral adipose tissue strongly relates to the manifestations of psoriasis. Skin and adipose tissue interact through adipokines. One of these adipokines—adiponectin—is a well-known regulator of insulin sensitivity, which also modulates proliferation and migration of keratinocytes [28] and modifies psoriasiform skin inflammation through suppression of IL-17 production [29]. Consistent with these observations, adiponectin KO mice can demonstrate a severe form of psoriasiform dermatitis [29]. On the other hand, HFD significantly exacerbates early psoriatic skin inflammation independent of obesity. Furthermore, a reduction of dietary saturated fatty acids improves psoriasiform inflammation independently of weight and fat content [30].
We have proposed that it is not the abdominal adipose tissue; rather, it is the dWAT located at the interface between dermis/subcutis that is involved in the pathophysiology of psoriasis [31,32]. There are several direct indications that dWAT undergoes structural modifications strongly associated with the psoriatic phenotype. PUVA treatment (a combination treatment consisting of application of psoralens (light sensitizers) (P) and then exposing the skin to UVA (long wave ultraviolet radiation) of K5.hTGFb1 transgenic mice (a murine model for psoriasis) caused a significant reduction of the dWAT layer beneath the psoriatic lesions, and such a modification correlated with clinical skin improvements in these animals [33]. Sonoelastographic investigation of the fat tissue located beneath the human psoriatic plaques demonstrates that the mechanical properties of the subcutis, beneath these spatially restricted areas, are very different from those in the areas covered by healthy skin. Successful treatment of psoriasis leads to skin improvements and simultaneous normalization of the adjacent fat tissue [34,35].
Taking into account the spatially limited distribution of psoriatic lesions and the modifications of dWAT located beneath these lesions [31,32], a critical question is what happens with dWAT during exposure to HFD? As we have demonstrated recently, HFD prompts at least two major modifications to dWAT: (1) a dramatic expansion of its volume involving both the processes of hyperplasia and hypertrophy of dermal adipocytes, and (2) a partial synchronization of HFs in the catagen/telogen phases with an impairment of their re-entry into anagen compared to chow-fed controls [4] ( ). Both modifications effectively increase the number of spatially and temporally correlated de-differentiation and re-differentiation events that mature adipocytes undergo. Consequently, the relative levels of lipids in skin in the catagen stage of the HF cycle, as well as the production of the antibacterial cathelicidin protein in the anagen, differ. Of note, such dWAT modifications also modulate the mechanical properties of the skin. Figure 2. Influence of the high-fat diet on the dWAT structure and HF cycle [4] (with permission of The Journal of Clinical Investigation).
Influence of the high-fat diet on the dWAT structure and HF cycle [4] (with permission of The Journal of Clinical Investigation).
Many of these pathological changes discussed here may be relevant to other skin diseases beyond Acne and Psoriasis. This includes hidradenitis suppurativa and other pathological alterations in the skin. Future experimental approaches will have to put the underlying mechanistic connections that we propose here into the test.