Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Alison Ramser and Version 2 by Dean Liu.
Originally named for its expression in the posterior hypothalamus in rats and after the Greek word for “appetite”, hypocretin, or orexin, as it is known today, gained notoriety as a neuropeptide regulating feeding behavior, energy homeostasis, and sleep. Orexin is a neuropeptide involved in both central and peripheral control of neuroendocrine functions, energy balance, and metabolism.
- orexin
- central regulation
- peripheral regulation
- metabolism
1. Overview of Orexin
Genomic research during the 1990s resulted in the characterization of numerous, previously unidentified, genes with potential biological significance. Notably, several “orphan” G-protein coupled receptors were found and shown to be putative, but without known ligands [1][10]. Given that G-protein coupled receptors are the most targeted molecules for drugs used in clinics, the investigation into the ligands for these orphan receptors was undertaken and revealed two peptide ligands termed “orexins”, also known as hypocretin [1][2][10,11]. It has been shown that the administration of orexins into the central nervous system (CNS) resulted in increased food intake in mice and rats, and that their production was dependent on the nutritional state. Additionally, a group of neurons located in the lateral hypothalamic area of the brain produce the neuropeptide called orexin in two forms, orexin A and orexin B [1][2][10,11]. These forms are synthesized from proteolytic cleavage of the precursor, prepro-orexin. Orexin-A is composed of 33 amino acids with an N-terminal pyroglutamyl residue and a C-terminal amidation [3][4][12,13]. Its structure consists of four Cysteine residues forming sets of intra-chain disulfide bonds. This structure is conserved across mammalian species, including rats, mice, cows, sheep, pigs, and dogs [4][13]. Orexin-B is made up of 28 amino acid residues, and while its C-terminal is similar to orexin-A, its N-terminal is more variable [1][10]. Additionally, there are more differences in orexin-B amino acid sequences across mammalian species and chicken [5][14].
There are two known orexin receptors, orexin-1 receptor (OX1R) and orexin-2 receptor (OX2R). These are G-protein coupled receptors with OX1R being structurally similar to neuropeptide receptors such as the Y2 neuropeptide Y (NPY) receptor and thyrotropin-releasing hormone (TRH) receptor [1][6][7][10,15,16]. Both receptor genes are highly conserved across species [8][17]. Orexin-A has a higher affinity for OX1R than orexin-B, while OX2R is a nonselective receptor for both orexin-A and orexin-B [1][3][10,12]. This difference in affinity for OX1R is attributed to the N-terminal of orexin-A, which is specific and hydrophilic [9][18]. Within the CNS, OX1R is the most abundant in the locus coeruleus, but is also found in the prefrontal and infralimbic cortex, hippocampus, amygdala, periventricular nucleus, anterior hypothalamus, dorsal raphe nucleus, and laterodorsal tegmental nucleus [10][11][4,19]. OX2R is expressed in the amygdala periventricular nucleus, dorsal raphe nucleus, and laterodorsal tegmental nucleus, as well as in the tuberomammillary nucleus. These regions are critical for energy homeostasis responses and arousal, which will be discussed in more detail in a later section. Furthermore, the mRNA of prepro-orexin is expressed in the lateral hypothalamus area, known as a feeding center [10][12][4,9]. The orexin system is also expressed in peripheral tissues, including the kidney, adrenal gland, thyroid, testes, ovaries, jejunum, lung, pituitary gland, brown and white adipose tissues, and muscle [13][14][5][15][16][17][18][19][6,7,14,20,21,22,23,24].
2. Central Orexin Signaling Pathways
The molecular mechanisms of central orexin’s mode of action have also been investigated. In energy restricted dairy cows, orexin A neurons are colocalized with adenosine monophosphate-activated protein kinase (AMPK) and peroxisome proliferator-activated receptor (PPAR) gamma. Additionally, energy restriction phosphorylates AMPK, leading to AMPK’s activation and to an increased PPARγ expression, indicating that orexin-A’s control of energy homeostasis in dairy cows involves AMPK [20][44]. AMPK is an enzyme that acts as a sensor for cellular energy status through changes in the ATP-to-AMP ratio and activating downstream targets to induce metabolic shifts [21][22][45,46]. AMPK is known to affect fat metabolism and glucose utilization, as well as impact the balance between catabolic and anabolic pathways within the cell [22][46]. PPARγ is a receptor whose ligands are known as potent insulin sensitizers, and it is known for its involvement in the mobilization of lipids and glucose metabolism [23][47]. It was later shown that orexin-A activated hypothalamic AMPK signaling in a calcium-dependent manner via a voltage-gated L-type calcium channel (Figure 12) [24][48]. This research points to orexins’ ability to directly activate AMPK signaling within the central nervous system, providing a means in which feeding behavior and energy homeostasis are linked. A link between central regulation and energy homeostasis was also established via hypothalamic orexin expression and brown adipose tissue thermogenesis. AMPK inhibition in the ventromedial nucleus of the hypothalamus, followed by increased orexin signaling in the lateral hypothalamic area, was seen under thermogenic effects induced by bone morphogenetic protein (BMP) 8B in mice. The thermogenic effect of BMP8B is due to its impact on the browning of white adipose tissue, and both its thermogenic effect and the effects on orexin expression were reduced by the knockout of glutamate vesicular transporter 2 (VGLUT2) [25][49]. These findings show the central control of energy homeostasis via orexin’s relationship to AMPK within the hypothalamus.
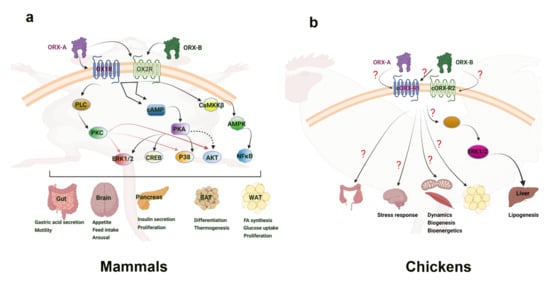
Figure 12. Orexin signaling pathways in mammalian (a) and avian species (b). “?” means that the downstream mediators are still not known or not well defined. Clear connections have been made to the link between orexin and the activation of energy sensing AMPK within the central nervous system of mammals. This provides a basis for connecting feeding behavior with orexin. Additionally, the ERK1/2 and Akt signaling pathways have been seen in hepatic response to orexin, inducing diverse cellular responses. While the main mechanism of action has yet to be determined, adipose tissue is shown to increase cytokines and other factors in response to orexin. In avian species, the ERK1/2 pathway has also been implicated in the liver response to orexin. Several mitochondrial related genes were shown to respond to orexin in avian muscle, but the direct mechanism of impact has yet to be elucidated. Culpable molecular signaling pathways have yet to be discovered in the avian central nervous system and adipose tissue response to orexin. AKT—Ak strain transforming kinase; AMPK—adenosine monophosphate-activated protein kinase; BAT—brown adipose tissue; CaMKKβ—Calcium/Calmodulin dependent protein kinase; cAMP—cyclic adenosine monophosphate; cORX-R1; chicken orexin receptor 1; cORX-R2—chicken orexin receptor 2; CREB—cAMP-response element binding protein; ERK1/2—extracellular-signal-regulated kinase 1/2; NFκB—nuclear factor kappa light chain enhancer of activated B cells; ORX-A—orexin-A; ORX-B—orexin-B; OXR1—orexin receptor 1; OXR2—orexin receptor 2; PKA—protein kinase A; PKC—protein kinase C; PLC—phospholipase C; P38—mitogen-associated protein kinase 38; WAT—white adipose tissue.
3. Orexin in the Avian Central Nervous System
The central role of orexins in avian species is a stark contrast to mammalian species. In fact, i.c.v. administration of orexins in neonatal chicks failed to stimulate appetite and food intake [26][50]. This is despite the fact that orexin-A and -B are highly conserved among vertebrates, as evidenced by the predicted amino-acid sequence of chicken prepro-orexin [27][51]. Additionally, while orexin-positive cell bodies were found in the periventricular hypothalamic nucleus and extending into the lateral hypothalamic area in chicken, fasting had no effect on the orexin mRNA expression [27][51]. These phenomena were further investigated upon research into the chicken orexin receptor (cOXR). It was found that cOXR corresponds more closely to OX2R in mammals, with approximately 80% similarity [5][14]. cOXR was found to be widely expressed throughout the bird brain, and particularly abundant in the cerebrum, hypothalamus, and optic tectum [5][14]. It was later found that orexin-A and -B do not seem to be involved in the wake-sleep cycle, as their expression levels did not change in the brains of sleeping vs. awake laying hens [28][52]. Within the avian brain, orexin-A and -B neurons have been found central on the paraventricular nucleus and extending into the lateral hypothalamic area in several birds [29][30][53,54]. The highest density of orexin neurons in the house finch was found within the preoptic area, the hypothalamus, and the thalamus, with projections also found in the third ventricle caudally [30][54]. The distribution of orexin neurons in varying regions within the avian brain point to a diverse involvement of orexin in numerous behavioral and centrally-regulated functions. A recent study by Wei and colleagues showed that keel fracture induced stress and inflammation, along with a reduced expression of the orexin system in the hypothalamus of laying hens [31][55]. This would implicate a central expression of orexin in stress responses, similar to what was seen in mammalian species. On the other hand, Lei et al. reported that acute heat stress did not elicit any change to the hypothalamic expression of orexin mRNA in broiler chickens [32][56]. Taken together, these studies suggest a potential CNS role for the avian orexin system in certain stress responses, but not in energy balance, and this role may be strain-dependent (layers vs. broilers). Therefore, further in-depth investigations are needed to elucidate the mechanisms in which orexin operates within the avian CNS and its downstream effects.